|
Biodiversity Data Journal :
Species inventory
|
Spatial distribution of Madeira Island Laurisilva endemic spiders (Arachnida: Araneae)
|
Corresponding author:
Academic editor: Emma McCarroll Shaw
Received: 07 Jan 2014 | Accepted: 12 Feb 2014 | Published: 14 Feb 2014
© 2014 Luís Crespo, Mário Boieiro, Pedro Cardoso, Carlos Aguiar, Isabel Amorim, Carla Barrinha, Paulo Borges, Dília Menezes, Fernando Pereira, Carla Rego, Sérvio Ribeiro, Israel Silva, Artur Serrano
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Crespo L, Boieiro M, Cardoso P, Aguiar C, Amorim I, Barrinha C, Borges P, Menezes D, Pereira F, Rego C, Ribeiro S, Silva I, Serrano A (2014) Spatial distribution of Madeira Island Laurisilva endemic spiders (Arachnida: Araneae). Biodiversity Data Journal 2: e1051. https://doi.org/10.3897/BDJ.2.e1051
|
|
Abstract
Madeira island presents a unique spider diversity with a high number of endemic species, many of which are still poorly known. A recent biodiversity survey on the terrestrial arthropods of the native forest, Laurisilva, provided a large set of standardized samples from various patches throughout the island. Out of the fifty two species recorded, approximately 33.3% are Madeiran endemics, many of which had not been collected since their original description. Two new species to science are reported – Ceratinopsis n. sp. and Theridion n. sp. – and the first records of Poeciloneta variegata (Blackwall, 1841) and Tetragnatha intermedia Kulczynski, 1891 are reported for the first time for Madeira island. Considerations on species richness and abundance from different Laurisilva locations are presented, together with distribution maps for endemic species. These results contribute to a better understanding of spider diversity patterns and endemic species distribution in the native forest of Madeira island.
Keywords
Macaronesia, terrestrial arthropods, island endemics, laurel forest, biodiversity hotspot
Introduction
Interest on Madeira spider fauna started almost two centuries ago (
Laurisilva is a relict evergreen forest that covered part of the Mediterranean Basin during the Tertiary, being restricted nowadays to a few locations in the Macaronesian archipelagoes of the Azores, Madeira and Canary Islands, and in some secluded sites of North Africa (
The latest spider checklist (
Materials and methods
Spiders were sampled using two standardized complementary techniques: pitfall trapping and direct sampling. Pitfall sampling was performed both in May-June 2006 (sites 1 to 28) and June 2007 (sites 29 to 36). A linear transect with 30 pitfall traps, 5 m apart, was set in each study site. Each trap consisted of a plastic cup (4.2 cm diameter by 7.8 cm height) protected with a plastic cover (15 cm diameter) attached ~3 cm aboveground to prevent flooding and loss of specimens. Ethyleneglicol (10%) and Turquin solution were used as preservatives in alternate traps along the linear transect (BALA protocol,
Direct sampling was performed in June 2007 for most sampling sites. The sampling design was semi-quantitative with a sampling unit defined as 30 minute/person of effective fieldwork. Direct sampling included the search of spiders in three microhabitats (under bark of dead tree trunks, mosses and lichens, on living tree trunks and under stones/leaf litter) with the collection of six replicates/microhabitat/site (total 18 samples per site).
For each spider species collected, we recorded information on sampling technique used, site location code (Table
Study sites, sampling date/period and collecting method. Geographic coordinates in decimal degrees.
| Study sites | Sampling | |||||
|---|---|---|---|---|---|---|
| Code | Site | Lat. (N) | Long. (W) | Altitude (m) | Pitfall trapping period | Direct sampling period |
| 1 | Funduras | 32.7493 | -16.8114 | 500 | 22.V-05.VI.2006 | 04.VI.2007 |
| 2 | Funduras | 32.7540 | -16.8099 | 552 | 22.V-05.VI.2006 | 04.VI.2007 |
| 3 | Fajã da Nogueira – Levada Pte. Roquete | 32.7391 | -16.9156 | 1074 | 23.V-06.VI.2006 | - |
| 4 | Fajã da Nogueira – Mtdo. do Leacoque | 32.7415 | -16.9161 | 630 | 23.V-06.VI.2006 | 06.VI.2007 |
| 5 | Fajã da Nogueira – Casa do Levadeiro | 32.7406 | -16.9136 | 989 | 23.V-06.VI.2006 | - |
| 6 | Fajã da Nogueira – Mtdo. do Leacoque | 32.7418 | -16.9177 | 614 | 23.V-06.VI.2006 | 06.VI.2007 |
| 7 | Fajã da Nogueira – Tanque | 32.7425 | -16.9168 | 845 | 23.V-06.VI.2006 | 06.VI.2007 |
| 8 | Fajã da Nogueira – Til Gigante | 32.7457 | -16.9150 | 841 | 23.V-06.VI.2006 | 06.VI.2007 |
| 9 | Queimadas | 32.7873 | -16.9047 | 841 | 24.V-07.VI.2006 | 05.VI.2007 |
| 10 | Pico das Pedras | 32.7841 | -16.9055 | 883 | 24.V-07.VI.2006 | 05.VI.2007 |
| 11 | Achada do Teixeira | 32.7733 | -16.9081 | 1211 | 24.V-07.VI.2006 | 05.VI.2007 |
| 12 | Achada do Teixeira | 32.7762 | -16.9022 | 1103 | 24.V-07.VI.2006 | 05.VI.2007 |
| 13 | Ribeiro Frio – Viveiro | 32.7354 | -16.8864 | 906 | 24.V-06.VI.2006 | 05.VI.2007 |
| 14 | Ribeiro Frio – Cottages | 32.7319 | -16.8861 | 994 | 24.V-07.VI.2006 | 06.VI.2007 |
| 15 | Pico do Areeiro | 32.7231 | -16.9109 | 1533 | 24.V-07.VI.2006 | - |
| 16 | Pico do Areeiro | 32.7287 | -16.9202 | 1594 | 24.V-07.VI.2006 | - |
| 17 | Chão da Ribeira | 32.7933 | -17.1122 | 519 | 25.V-08.VI.2006 | 07.VI.2007 |
| 18 | Chão da Ribeira | 32.7957 | -17.1117 | 491 | 25.V-08.VI.2006 | 07.VI.2007 |
| 19 | Chão dos Louros | 32.7636 | -17.0190 | 748 | 25.V-08.VI.2006 | 07.VI.2007 |
| 20 | Encumeada | 32.7558 | -17.0143 | 999 | 25.V-08.VI.2006 | - |
| 21 | Ribeiro Bonito – Levada | 32.8047 | -16.9346 | 568 | 25.V-08.VI.2006 | 09.VI.2007 |
| 22 | Ribeiro Bonito – Ribeiro | 32.7985 | -16.9360 | 560 | 25.V-08.VI.2006 | 09.VI.2007 |
| 23 | Fanal | 32.8302 | -17.1585 | 755 | 26.V-09.VI.2006 | - |
| 24 | Fanal – Levada dos Cedros | 32.8259 | -17.1580 | 820 | 26.V-09.VI.2006 | 09.VI.2007 |
| 25 | Fanal | 32.8236 | -17.1560 | 890 | 26.V-09.VI.2006 | 09.VI.2007 |
| 26 | Fanal | 32.8226 | -17.1539 | 889 | 26.V-09.VI.2006 | 09.VI.2007 |
| 27 | Fanal | 32.8182 | -17.1521 | 1023 | 26.V-09.VI.2006 | 09.VI.2007 |
| 28 | Fanal | 32.8062 | -17.1409 | 1134 | 26.V-09.VI.2006 | 10.VI.2007 |
| 29 | Ginjas | 32.7758 | -17.0534 | 869 | 08-22.VI.2007 | 23.VI.2007 |
| 30 | Caramujo | 32.7722 | -17.0529 | 981 | 08-22.VI.2007 | - |
| 31 | Caramujo | 32.7746 | -17.0559 | 1001 | 08-22.VI.2007 | - |
| 32 | Rabaças | 32.7413 | -17.0783 | 993 | 10-24.VI.2007 | - |
| 33 | Rabaçal | 32.7647 | -17.1341 | 930 | 10-24.VI.2007 | 25.VI.2007 |
| 34 | Risco | 32.7608 | -17.1256 | 1048 | 10-24.VI.2007 | 25.VI.2007 |
| 35 | Casa do Elias | 32.8268 | -17.1883 | 814 | 26.VI-10.VII.2007 | - |
| 36 | Galhano | 32.7971 | -17.1729 | 975 | 27.VI-11.VII.2007 | - |
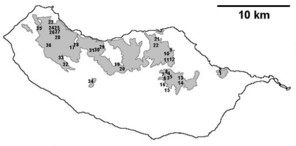
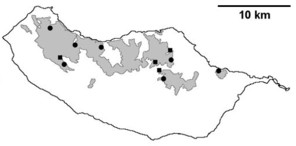
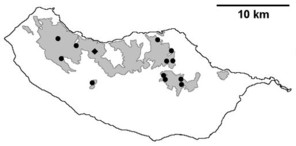
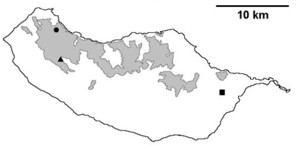
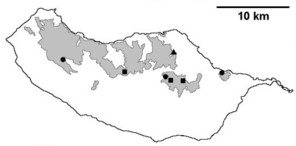
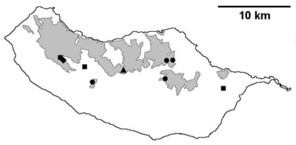
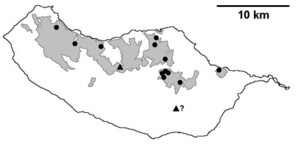
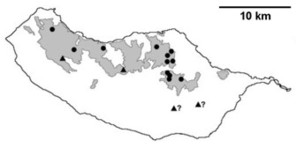
Sampling sites and distribution of endemic spider species in Madeira island. See Table
b: distribution of Macarophaeus cultior;
c: distribution of Hahnia insulana;
d: distribution of Centromerus variegatus.
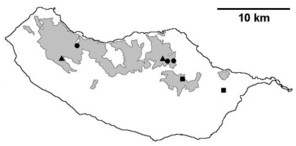
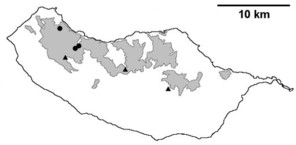
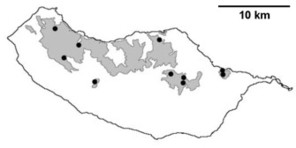
Distribution of endemic spider species in Madeira island. Circles: present records; rhombuses: records from
b: Ceratinopsis infuscata
c: Ceratinopsis n. sp.
d: Frontinellina dearmata
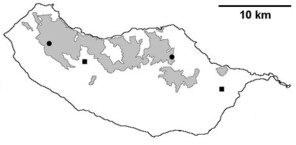
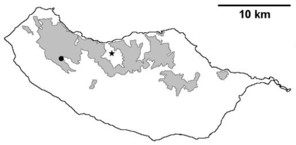
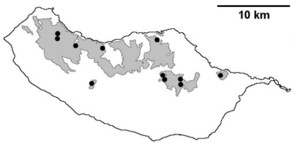
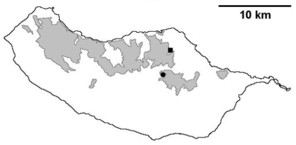
Distribution of endemic spider species in Madeira island. Circles: present records; rhombuses: records from
b: Lepthyphantes impudicus;
c: Lepthyphantes mauli;
d: Turinyphia maderiana.
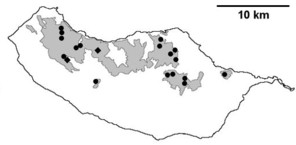
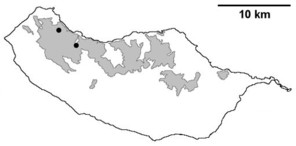
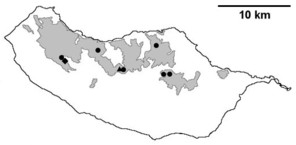
Distribution of endemic spider species in Madeira island. Circles: present records; rhombuses: records from
b: Meta stridulans;
c: Dipoenata longitarsis;
d: Episinus maderianus.
Faunistic results
Laurisilva of Madeira
The study took place in Madeira island located in the Atlantic Ocean, approximately 1000 km from the Iberian Peninsula. Sampling was conducted in thirty-six sites (Table




Zygiella minima
-
locationID: 33; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Rabaçal; verbatimElevation:930; decimalLatitude:32.7647; decimalLongitude:-17.1341; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
This species was recently found for the first time in Madeira archipelago in an erosion cave of Deserta Grande island (
Clubiona decora
-
locationID: 1; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:500; decimalLatitude:32.7493; decimalLongitude:-16.8114; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 6; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:614; decimalLatitude:32.7418; decimalLongitude:-16.9177; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 7; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Tanque; verbatimElevation:845; decimalLatitude:32.7425; decimalLongitude:-16.9168; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 11; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1211; decimalLatitude:32.7733; decimalLongitude:-16.9081; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 2 females
-
locationID: 24; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal - Levada dos Cedros; verbatimElevation:820; decimalLatitude:32.8259; decimalLongitude:-17.158; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 25; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:890; decimalLatitude:32.8236; decimalLongitude:-17.156; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 33; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Rabaçal; verbatimElevation:930; decimalLatitude:32.7647; decimalLongitude:-17.1341; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
Its presence in laurel forest suggests being a native species and its occurence in the Balkans needs to be confirmed. In the Azores the species is very common in the laurel forest and in the canopies of low altitude orchards and exotic trees.
Lathys affinis
-
locationID: 4; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:630; decimalLatitude:32.7415; decimalLongitude:-16.9161; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 6; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:614; decimalLatitude:32.7418; decimalLongitude:-16.9177; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 7; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Tanque; verbatimElevation:845; decimalLatitude:32.7425; decimalLongitude:-16.9168; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 11; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1211; decimalLatitude:32.7733; decimalLongitude:-16.9081; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 4 females
-
locationID: 12; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1103; decimalLatitude:32.7762; decimalLongitude:-16.9022; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 13; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Viveiro; verbatimElevation:906; decimalLatitude:32.7354; decimalLongitude:-16.8864; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 2 females
-
locationID: 14; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Cottages; verbatimElevation:994; decimalLatitude:32.7319; decimalLongitude:-16.8861; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:5 females
-
locationID: 24; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal - Levada dos Cedros; verbatimElevation:820; decimalLatitude:32.8259; decimalLongitude:-17.158; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:4 females
-
locationID: 24; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal - Levada dos Cedros; verbatimElevation:820; decimalLatitude:32.8259; decimalLongitude:-17.158; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male
-
locationID: 25; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:890; decimalLatitude:32.8236; decimalLongitude:-17.156; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 27; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1023; decimalLatitude:32.8182; decimalLongitude:-17.1521; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 28; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1134; decimalLatitude:32.8062; decimalLongitude:-17.1409; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:3 females
-
locationID: 33; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Rabaçal; verbatimElevation:930; decimalLatitude:32.7647; decimalLongitude:-17.1341; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:3 females
-
locationID: 34; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Ponta do Sol; locality:Risco; verbatimElevation:1048; decimalLatitude:32.7608; decimalLongitude:-17.1256; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
This macaronesian endemic is widespread in the Laurisilva of Madeira island. L. Crespo has examined the material cited as L. affinis from the Canary Islands, deposited at the Senckenberg Naturmuseum of Frankfurt, and questions their species affiliation. Additional taxonomic analyses are thus needed to clarify the taxonomic status of the Canary Islands specimens.
Nigma puella
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
Haplodrassus dalmatensis
-
locationID: 15; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1533; decimalLatitude:32.7231; decimalLongitude:-16.9109; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:13 males, 4 females
-
locationID: 16; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1594; decimalLatitude:32.7287; decimalLongitude:-16.9202; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:20 males, 19 females
Macarophaeus cultior
-
locationID: 2; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:552; decimalLatitude:32.754; decimalLongitude:-16.8099; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 6; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:614; decimalLatitude:32.7418; decimalLongitude:-16.9177; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 5 females
-
locationID: 12; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1103; decimalLatitude:32.7762; decimalLongitude:-16.9022; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 1 female
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 2 females
-
locationID: 24; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal - Levada dos Cedros; verbatimElevation:820; decimalLatitude:32.8259; decimalLongitude:-17.158; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 3 females
-
locationID: 29; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Ginjas; verbatimElevation:869; decimalLatitude:32.7758; decimalLongitude:-17.0534; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 2 females
Madeira island (Fig.
Micaria pallipes
-
locationID: 15; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1533; decimalLatitude:32.7231; decimalLongitude:-16.9109; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:9 males, 2 females
-
locationID: 16; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1594; decimalLatitude:32.7287; decimalLongitude:-16.9202; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:6 males, 7 females
Zelotes civicus
-
locationID: 15; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1533; decimalLatitude:32.7231; decimalLongitude:-16.9109; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:54 males, 25 females
-
locationID: 16; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1594; decimalLatitude:32.7287; decimalLongitude:-16.9202; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:99 males, 36 females
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:3 males
Hahnia insulana
-
locationID: 6; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:614; decimalLatitude:32.7418; decimalLongitude:-16.9177; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 7; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Tanque; verbatimElevation:845; decimalLatitude:32.7425; decimalLongitude:-16.9168; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 males
-
locationID: 9; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Queimadas; verbatimElevation:841; decimalLatitude:32.7873; decimalLongitude:-16.9047; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 12; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1103; decimalLatitude:32.7762; decimalLongitude:-16.9022; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:3 males, 1 female
-
locationID: 13; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Viveiro; verbatimElevation:906; decimalLatitude:32.7354; decimalLongitude:-16.8864; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 14; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Cottages; verbatimElevation:994; decimalLatitude:32.7319; decimalLongitude:-16.8861; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 males
-
locationID: 21; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Levada; verbatimElevation:568; decimalLatitude:32.8047; decimalLongitude:-16.9346; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 33; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Rabaçal; verbatimElevation:930; decimalLatitude:32.7647; decimalLongitude:-17.1341; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 34; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Ponta do Sol; locality:Risco; verbatimElevation:1048; decimalLatitude:32.7608; decimalLongitude:-17.1256; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
Madeira island (Fig.
This is the first time this species was captured after its description (
Centromerus variegatus
-
locationID: 25; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:890; decimalLatitude:32.8236; decimalLongitude:-17.156; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 2 females
Madeira island (Fig.
Records of C. variegatus are extremely scarce. After its formal description based on five specimens collected in Santo da Serra (
Ceratinopsis acripes
-
locationID: 2; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:552; decimalLatitude:32.754; decimalLongitude:-16.8099; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 7; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Tanque; verbatimElevation:845; decimalLatitude:32.7425; decimalLongitude:-16.9168; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 33; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Rabaçal; verbatimElevation:930; decimalLatitude:32.7647; decimalLongitude:-17.1341; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
Madeira island (Fig.
Ceratinopsis acripes seems to be restricted to Laurisilva.
Ceratinopsis infuscata
-
locationID: 11; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1211; decimalLatitude:32.7733; decimalLongitude:-16.9081; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 12; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1103; decimalLatitude:32.7762; decimalLongitude:-16.9022; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
Madeira island (Fig.
Ceratinopsis infuscata seems to be restricted to Laurisilva, with an additional record from Santo da Serra (
Ceratinopsis n. sp.
-
locationID: 1; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:500; decimalLatitude:32.7493; decimalLongitude:-16.8114; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 6; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:614; decimalLatitude:32.7418; decimalLongitude:-16.9177; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 7; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Tanque; verbatimElevation:845; decimalLatitude:32.7425; decimalLongitude:-16.9168; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 13; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Viveiro; verbatimElevation:906; decimalLatitude:32.7354; decimalLongitude:-16.8864; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 2 females
-
locationID: 14; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Cottages; verbatimElevation:994; decimalLatitude:32.7319; decimalLongitude:-16.8861; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 1 female
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 2 females
-
locationID: 21; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Levada; verbatimElevation:568; decimalLatitude:32.8047; decimalLongitude:-16.9346; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 27; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1023; decimalLatitude:32.8182; decimalLongitude:-17.1521; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 28; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1134; decimalLatitude:32.8062; decimalLongitude:-17.1409; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 29; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Ginjas; verbatimElevation:869; decimalLatitude:32.7758; decimalLongitude:-17.0534; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 34; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Ponta do Sol; locality:Risco; verbatimElevation:1048; decimalLatitude:32.7608; decimalLongitude:-17.1256; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
Madeira island (Fig.
This is a new species to science, to be described in a future work. As the previous species of Ceratinopsis, it also seems to be restricted to Laurisilva.
Diplostyla concolor
-
locationID: 3; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Pte. Roquete; verbatimElevation:1074; decimalLatitude:32.7391; decimalLongitude:-16.9156; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 16; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1594; decimalLatitude:32.7287; decimalLongitude:-16.9202; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 females
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 20; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Encumeada; verbatimElevation:999; decimalLatitude:32.7558; decimalLongitude:-17.0143; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
Entelecara schmitzi
-
locationID: 6; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:614; decimalLatitude:32.7418; decimalLongitude:-16.9177; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 7; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Tanque; verbatimElevation:845; decimalLatitude:32.7425; decimalLongitude:-16.9168; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 8; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Til Gigante; verbatimElevation:841; decimalLatitude:32.7457; decimalLongitude:-16.915; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 11; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1211; decimalLatitude:32.7733; decimalLongitude:-16.9081; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 1 female
-
locationID: 13; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Viveiro; verbatimElevation:906; decimalLatitude:32.7354; decimalLongitude:-16.8864; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 14; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Cottages; verbatimElevation:994; decimalLatitude:32.7319; decimalLongitude:-16.8861; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 17; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:519; decimalLatitude:32.7933; decimalLongitude:-17.1122; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 24; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal - Levada dos Cedros; verbatimElevation:820; decimalLatitude:32.8259; decimalLongitude:-17.158; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:8 females
-
locationID: 25; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:890; decimalLatitude:32.8236; decimalLongitude:-17.156; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 26; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:889; decimalLatitude:32.8226; decimalLongitude:-17.1539; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 2 females
-
locationID: 27; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1023; decimalLatitude:32.8182; decimalLongitude:-17.1521; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 29; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Ginjas; verbatimElevation:869; decimalLatitude:32.7758; decimalLongitude:-17.0534; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 34; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Ponta do Sol; locality:Risco; verbatimElevation:1048; decimalLatitude:32.7608; decimalLongitude:-17.1256; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
Frontinellina dearmata
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 25; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:890; decimalLatitude:32.8236; decimalLongitude:-17.156; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 males
Madeira island (Fig.
Frontiphantes fulgurenotatus
-
locationID: 6; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:614; decimalLatitude:32.7418; decimalLongitude:-16.9177; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 males
-
locationID: 11; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1211; decimalLatitude:32.7733; decimalLongitude:-16.9081; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 12; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1103; decimalLatitude:32.7762; decimalLongitude:-16.9022; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 34; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Ponta do Sol; locality:Risco; verbatimElevation:1048; decimalLatitude:32.7608; decimalLongitude:-17.1256; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
Madeira island (Fig.
This endemic species seems to be restricted to Laurisilva. The two records outside Laurisilva (
Lepthyphantes impudicus
-
locationID: 12; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1103; decimalLatitude:32.7762; decimalLongitude:-16.9022; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 36; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Galhano; verbatimElevation:975; decimalLatitude:32.7971; decimalLongitude:-17.1729; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
Madeira island (Fig.
Distribution data on L. impudicus is scarce consisting of only 4 records. This species seems to be associated with Laurisilva and the lack of congruence between historical data (
Lepthyphantes mauli
-
locationID: 33; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Rabaçal; verbatimElevation:930; decimalLatitude:32.7647; decimalLongitude:-17.1341; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male
Madeira island (Fig.
Meioneta fuscipalpa
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 males
Microctenonyx subitaneus
-
locationID: 16; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1594; decimalLatitude:32.7287; decimalLongitude:-16.9202; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
Microlinyphia johnsoni
-
locationID: 8; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Til Gigante; verbatimElevation:841; decimalLatitude:32.7457; decimalLongitude:-16.915; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
Palliduphantes schmitzi
-
locationID: 1; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:500; decimalLatitude:32.7493; decimalLongitude:-16.8114; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 females
-
locationID: 2; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:552; decimalLatitude:32.754; decimalLongitude:-16.8099; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 males, 8 females
-
locationID: 3; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Pte. Roquete; verbatimElevation:1074; decimalLatitude:32.7391; decimalLongitude:-16.9156; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 4; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:630; decimalLatitude:32.7415; decimalLongitude:-16.9161; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:4 females
-
locationID: 4; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:630; decimalLatitude:32.7415; decimalLongitude:-16.9161; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 5; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Casa do Levadeiro; verbatimElevation:989; decimalLatitude:32.7406; decimalLongitude:-16.9136; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 2 females
-
locationID: 6; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:614; decimalLatitude:32.7418; decimalLongitude:-16.9177; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 7; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Tanque; verbatimElevation:845; decimalLatitude:32.7425; decimalLongitude:-16.9168; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 1 female
-
locationID: 9; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Queimadas; verbatimElevation:841; decimalLatitude:32.7873; decimalLongitude:-16.9047; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 1 female
-
locationID: 10; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico das Pedras; verbatimElevation:883; decimalLatitude:32.7841; decimalLongitude:-16.9055; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 10; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico das Pedras; verbatimElevation:883; decimalLatitude:32.7841; decimalLongitude:-16.9055; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 2 females
-
locationID: 11; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1211; decimalLatitude:32.7733; decimalLongitude:-16.9081; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 11; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1211; decimalLatitude:32.7733; decimalLongitude:-16.9081; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:4 males, 2 females
-
locationID: 12; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1103; decimalLatitude:32.7762; decimalLongitude:-16.9022; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 males, 3 females
-
locationID: 13; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Viveiro; verbatimElevation:906; decimalLatitude:32.7354; decimalLongitude:-16.8864; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 1 female
-
locationID: 15; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1533; decimalLatitude:32.7231; decimalLongitude:-16.9109; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male
-
locationID: 17; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:519; decimalLatitude:32.7933; decimalLongitude:-17.1122; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 males
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 2 females
-
locationID: 19; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Chão dos Louros; verbatimElevation:748; decimalLatitude:32.7636; decimalLongitude:-17.019; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:3 females
-
locationID: 20; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Encumeada; verbatimElevation:999; decimalLatitude:32.7558; decimalLongitude:-17.0143; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 males
-
locationID: 21; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Levada; verbatimElevation:568; decimalLatitude:32.8047; decimalLongitude:-16.9346; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 21; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Levada; verbatimElevation:568; decimalLatitude:32.8047; decimalLongitude:-16.9346; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 22; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Ribeiro; verbatimElevation:560; decimalLatitude:32.7985; decimalLongitude:-16.936; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 28; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1134; decimalLatitude:32.8062; decimalLongitude:-17.1409; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 1 female
-
locationID: 29; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Ginjas; verbatimElevation:869; decimalLatitude:32.7758; decimalLongitude:-17.0534; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male
-
locationID: 30; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Caramujo; verbatimElevation:981; decimalLatitude:32.7722; decimalLongitude:-17.0529; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 1 female
-
locationID: 31; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Caramujo; verbatimElevation:1001; decimalLatitude:32.7746; decimalLongitude:-17.0559; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male
-
locationID: 36; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Galhano; verbatimElevation:975; decimalLatitude:32.7971; decimalLongitude:-17.1729; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 males
Poeciloneta variegata
-
locationID: 24; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal - Levada dos Cedros; verbatimElevation:820; decimalLatitude:32.8259; decimalLongitude:-17.158; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 28; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1134; decimalLatitude:32.8062; decimalLongitude:-17.1409; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 1 female
Tenuiphantes tenuis
-
locationID: 1; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:500; decimalLatitude:32.7493; decimalLongitude:-16.8114; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:4 males, 5 females
-
locationID: 1; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:500; decimalLatitude:32.7493; decimalLongitude:-16.8114; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 3 females
-
locationID: 2; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:552; decimalLatitude:32.754; decimalLongitude:-16.8099; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:19 males, 22 females
-
locationID: 2; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:552; decimalLatitude:32.754; decimalLongitude:-16.8099; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:5 males, 7 females
-
locationID: 3; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Pte. Roquete; verbatimElevation:1074; decimalLatitude:32.7391; decimalLongitude:-16.9156; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 4; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:630; decimalLatitude:32.7415; decimalLongitude:-16.9161; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:5 males, 6 females
-
locationID: 4; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:630; decimalLatitude:32.7415; decimalLongitude:-16.9161; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 4 females
-
locationID: 5; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Casa do Levadeiro; verbatimElevation:989; decimalLatitude:32.7406; decimalLongitude:-16.9136; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 6; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:614; decimalLatitude:32.7418; decimalLongitude:-16.9177; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:8 males, 16 females
-
locationID: 6; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:614; decimalLatitude:32.7418; decimalLongitude:-16.9177; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 males, 4 females
-
locationID: 7; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Tanque; verbatimElevation:845; decimalLatitude:32.7425; decimalLongitude:-16.9168; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:3 males, 7 females
-
locationID: 7; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Tanque; verbatimElevation:845; decimalLatitude:32.7425; decimalLongitude:-16.9168; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:4 males, 1 female
-
locationID: 8; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Til Gigante; verbatimElevation:841; decimalLatitude:32.7457; decimalLongitude:-16.915; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:4 males, 9 females
-
locationID: 8; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Til Gigante; verbatimElevation:841; decimalLatitude:32.7457; decimalLongitude:-16.915; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 males, 3 females
-
locationID: 9; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Queimadas; verbatimElevation:841; decimalLatitude:32.7873; decimalLongitude:-16.9047; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 3 females
-
locationID: 9; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Queimadas; verbatimElevation:841; decimalLatitude:32.7873; decimalLongitude:-16.9047; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 females
-
locationID: 12; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1103; decimalLatitude:32.7762; decimalLongitude:-16.9022; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 males, 1 female
-
locationID: 13; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Viveiro; verbatimElevation:906; decimalLatitude:32.7354; decimalLongitude:-16.8864; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 14; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Cottages; verbatimElevation:994; decimalLatitude:32.7319; decimalLongitude:-16.8861; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 14; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Cottages; verbatimElevation:994; decimalLatitude:32.7319; decimalLongitude:-16.8861; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:3 males, 2 females
-
locationID: 16; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1594; decimalLatitude:32.7287; decimalLongitude:-16.9202; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 17; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:519; decimalLatitude:32.7933; decimalLongitude:-17.1122; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:5 males, 8 females
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 15 females
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:4 males, 1 female
-
locationID: 19; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Chão dos Louros; verbatimElevation:748; decimalLatitude:32.7636; decimalLongitude:-17.019; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 2 females
-
locationID: 20; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Encumeada; verbatimElevation:999; decimalLatitude:32.7558; decimalLongitude:-17.0143; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 males, 4 females
-
locationID: 21; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Levada; verbatimElevation:568; decimalLatitude:32.8047; decimalLongitude:-16.9346; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:6 males, 18 females
-
locationID: 21; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Levada; verbatimElevation:568; decimalLatitude:32.8047; decimalLongitude:-16.9346; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:4 females
-
locationID: 22; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Ribeiro; verbatimElevation:560; decimalLatitude:32.7985; decimalLongitude:-16.936; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 13 females
-
locationID: 22; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Ribeiro; verbatimElevation:560; decimalLatitude:32.7985; decimalLongitude:-16.936; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 males, 2 females
-
locationID: 23; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:755; decimalLatitude:32.8302; decimalLongitude:-17.1585; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 1 female
-
locationID: 24; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal - Levada dos Cedros; verbatimElevation:820; decimalLatitude:32.8259; decimalLongitude:-17.158; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 24; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal - Levada dos Cedros; verbatimElevation:820; decimalLatitude:32.8259; decimalLongitude:-17.158; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:9 males, 3 females
-
locationID: 25; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:890; decimalLatitude:32.8236; decimalLongitude:-17.156; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 3 females
-
locationID: 26; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:889; decimalLatitude:32.8226; decimalLongitude:-17.1539; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male
-
locationID: 27; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1023; decimalLatitude:32.8182; decimalLongitude:-17.1521; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 27; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1023; decimalLatitude:32.8182; decimalLongitude:-17.1521; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 males, 2 females
-
locationID: 28; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1134; decimalLatitude:32.8062; decimalLongitude:-17.1409; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 28; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1134; decimalLatitude:32.8062; decimalLongitude:-17.1409; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:4 males, 2 females
-
locationID: 29; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Ginjas; verbatimElevation:869; decimalLatitude:32.7758; decimalLongitude:-17.0534; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 males, 2 females
-
locationID: 29; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Ginjas; verbatimElevation:869; decimalLatitude:32.7758; decimalLongitude:-17.0534; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:3 females
-
locationID: 30; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Caramujo; verbatimElevation:981; decimalLatitude:32.7722; decimalLongitude:-17.0529; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 females
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:5 males, 13 females
-
locationID: 33; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Rabaçal; verbatimElevation:930; decimalLatitude:32.7647; decimalLongitude:-17.1341; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 males, 3 females
-
locationID: 33; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Rabaçal; verbatimElevation:930; decimalLatitude:32.7647; decimalLongitude:-17.1341; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:5 males, 2 females
-
locationID: 34; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Ponta do Sol; locality:Risco; verbatimElevation:1048; decimalLatitude:32.7608; decimalLongitude:-17.1256; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 males, 2 females
-
locationID: 34; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Ponta do Sol; locality:Risco; verbatimElevation:1048; decimalLatitude:32.7608; decimalLongitude:-17.1256; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:4 females
-
locationID: 35; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Casa do Elias; verbatimElevation:814; decimalLatitude:32.8268; decimalLongitude:-17.1883; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 2 females
-
locationID: 36; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Galhano; verbatimElevation:975; decimalLatitude:32.7971; decimalLongitude:-17.1729; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:6 males, 6 females
Turinyphia maderiana
-
locationID: 4; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:630; decimalLatitude:32.7415; decimalLongitude:-16.9161; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 1 female
-
locationID: 7; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Tanque; verbatimElevation:845; decimalLatitude:32.7425; decimalLongitude:-16.9168; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 20; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Encumeada; verbatimElevation:999; decimalLatitude:32.7558; decimalLongitude:-17.0143; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male
-
locationID: 22; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Ribeiro; verbatimElevation:560; decimalLatitude:32.7985; decimalLongitude:-16.936; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 30; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Caramujo; verbatimElevation:981; decimalLatitude:32.7722; decimalLongitude:-17.0529; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male
-
locationID: 33; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Rabaçal; verbatimElevation:930; decimalLatitude:32.7647; decimalLongitude:-17.1341; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
Madeira island (Fig.
This is the third record of this species. Turinyphia maderiana seems to be restricted to Laurisilva.
Typhochrestus madeirensis
-
locationID: 16; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1594; decimalLatitude:32.7287; decimalLongitude:-16.9202; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male
This species was recently described (
Unidentified species
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 1 female
Pardosa proxima
-
locationID: 15; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1533; decimalLatitude:32.7231; decimalLongitude:-16.9109; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:6 females
-
locationID: 16; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1594; decimalLatitude:32.7287; decimalLongitude:-16.9202; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:11 males, 51 females
Ero aphana
-
locationID: 2; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:552; decimalLatitude:32.754; decimalLongitude:-16.8099; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
Cheiracanthium albidulum
-
locationID: 2; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:552; decimalLatitude:32.754; decimalLongitude:-16.8099; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 6; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:614; decimalLatitude:32.7418; decimalLongitude:-16.9177; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 12; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1103; decimalLatitude:32.7762; decimalLongitude:-16.9022; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 13; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Viveiro; verbatimElevation:906; decimalLatitude:32.7354; decimalLongitude:-16.8864; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 24; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal - Levada dos Cedros; verbatimElevation:820; decimalLatitude:32.8259; decimalLongitude:-17.158; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:4 males, 1 female
-
locationID: 28; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1134; decimalLatitude:32.8062; decimalLongitude:-17.1409; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
Trogloneta madeirensis
-
locationID: 1; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:500; decimalLatitude:32.7493; decimalLongitude:-16.8114; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 4; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:630; decimalLatitude:32.7415; decimalLongitude:-16.9161; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:4 females
-
locationID: 6; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:614; decimalLatitude:32.7418; decimalLongitude:-16.9177; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 7; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Tanque; verbatimElevation:845; decimalLatitude:32.7425; decimalLongitude:-16.9168; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 2 females
-
locationID: 8; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Til Gigante; verbatimElevation:841; decimalLatitude:32.7457; decimalLongitude:-16.915; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 11; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1211; decimalLatitude:32.7733; decimalLongitude:-16.9081; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 14; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Cottages; verbatimElevation:994; decimalLatitude:32.7319; decimalLongitude:-16.8861; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:3 females
-
locationID: 21; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Levada; verbatimElevation:568; decimalLatitude:32.8047; decimalLongitude:-16.9346; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:4 males, 5 females
-
locationID: 22; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Ribeiro; verbatimElevation:560; decimalLatitude:32.7985; decimalLongitude:-16.936; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 1 female
-
locationID: 25; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:890; decimalLatitude:32.8236; decimalLongitude:-17.156; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 29; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Ginjas; verbatimElevation:869; decimalLatitude:32.7758; decimalLongitude:-17.0534; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
Madeira island (Fig.
This species seems to be restricted to Laurisilva. Wunderlich (
Philodromus insulanus
-
locationID: 28; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1134; decimalLatitude:32.8062; decimalLongitude:-17.1409; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
Thanatus oblongiusculus
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
Chalcoscirtus sublestus
-
locationID: 15; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1533; decimalLatitude:32.7231; decimalLongitude:-16.9109; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:5 males, 4 females
-
locationID: 16; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1594; decimalLatitude:32.7287; decimalLongitude:-16.9202; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 4 females
Macaroeris diligens
-
locationID: 4; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:630; decimalLatitude:32.7415; decimalLongitude:-16.9161; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 6; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:614; decimalLatitude:32.7418; decimalLongitude:-16.9177; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 11; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1211; decimalLatitude:32.7733; decimalLongitude:-16.9081; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 4 females
-
locationID: 12; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1103; decimalLatitude:32.7762; decimalLongitude:-16.9022; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 13; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Viveiro; verbatimElevation:906; decimalLatitude:32.7354; decimalLongitude:-16.8864; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 14; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Cottages; verbatimElevation:994; decimalLatitude:32.7319; decimalLongitude:-16.8861; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 24; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal - Levada dos Cedros; verbatimElevation:820; decimalLatitude:32.8259; decimalLongitude:-17.158; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:3 females
-
locationID: 25; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:890; decimalLatitude:32.8236; decimalLongitude:-17.156; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:4 females
-
locationID: 27; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1023; decimalLatitude:32.8182; decimalLongitude:-17.1521; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:3 females
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 33; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Rabaçal; verbatimElevation:930; decimalLatitude:32.7647; decimalLongitude:-17.1341; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 34; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Ponta do Sol; locality:Risco; verbatimElevation:1048; decimalLatitude:32.7608; decimalLongitude:-17.1256; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
Macaroeris sp.
-
locationID: 2; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:552; decimalLatitude:32.754; decimalLongitude:-16.8099; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 17; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:519; decimalLatitude:32.7933; decimalLongitude:-17.1122; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 males, 1 female
Pellenes maderianus
-
locationID: 15; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1533; decimalLatitude:32.7231; decimalLongitude:-16.9109; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male
Some confusion appears to exist regarding the identity of P. maderianus. Although most authors consider this species a valid taxon, some support that it is a synonymy of P. epularis (O. Pickard-Cambridge, 1872) (
Meta stridulans
-
locationID: 17; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:519; decimalLatitude:32.7933; decimalLongitude:-17.1122; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 25; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:890; decimalLatitude:32.8236; decimalLongitude:-17.156; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
Madeira island (Fig.
Tetragnatha intermedia
-
locationID: 2; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:552; decimalLatitude:32.754; decimalLongitude:-16.8099; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
First record for Madeira island. It is possible that previous records of T. obtusa C. L. Koch, 1837 from the Madeira archipelago (
Cryptachaea blattea
-
locationID: 1; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:500; decimalLatitude:32.7493; decimalLongitude:-16.8114; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 2 females
-
locationID: 2; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:552; decimalLatitude:32.754; decimalLongitude:-16.8099; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:4 females
-
locationID: 2; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:552; decimalLatitude:32.754; decimalLongitude:-16.8099; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 males, 2 females
-
locationID: 4; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:630; decimalLatitude:32.7415; decimalLongitude:-16.9161; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 6 females
-
locationID: 4; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:630; decimalLatitude:32.7415; decimalLongitude:-16.9161; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 females
-
locationID: 6; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:614; decimalLatitude:32.7418; decimalLongitude:-16.9177; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 6; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:614; decimalLatitude:32.7418; decimalLongitude:-16.9177; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 7; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Tanque; verbatimElevation:845; decimalLatitude:32.7425; decimalLongitude:-16.9168; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 males
-
locationID: 8; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Til Gigante; verbatimElevation:841; decimalLatitude:32.7457; decimalLongitude:-16.915; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 9; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Queimadas; verbatimElevation:841; decimalLatitude:32.7873; decimalLongitude:-16.9047; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 10; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico das Pedras; verbatimElevation:883; decimalLatitude:32.7841; decimalLongitude:-16.9055; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 4 females
-
locationID: 11; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1211; decimalLatitude:32.7733; decimalLongitude:-16.9081; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:4 males, 5 females
-
locationID: 11; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1211; decimalLatitude:32.7733; decimalLongitude:-16.9081; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 12; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1103; decimalLatitude:32.7762; decimalLongitude:-16.9022; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 males, 1 female
-
locationID: 12; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1103; decimalLatitude:32.7762; decimalLongitude:-16.9022; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 females
-
locationID: 13; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Viveiro; verbatimElevation:906; decimalLatitude:32.7354; decimalLongitude:-16.8864; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 males
-
locationID: 14; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Cottages; verbatimElevation:994; decimalLatitude:32.7319; decimalLongitude:-16.8861; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 15; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1533; decimalLatitude:32.7231; decimalLongitude:-16.9109; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 17; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:519; decimalLatitude:32.7933; decimalLongitude:-17.1122; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:3 females
-
locationID: 20; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Encumeada; verbatimElevation:999; decimalLatitude:32.7558; decimalLongitude:-17.0143; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 males, 4 females
-
locationID: 21; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Levada; verbatimElevation:568; decimalLatitude:32.8047; decimalLongitude:-16.9346; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:5 females
-
locationID: 21; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Levada; verbatimElevation:568; decimalLatitude:32.8047; decimalLongitude:-16.9346; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 22; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Ribeiro; verbatimElevation:560; decimalLatitude:32.7985; decimalLongitude:-16.936; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 22; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Ribeiro; verbatimElevation:560; decimalLatitude:32.7985; decimalLongitude:-16.936; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 23; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:755; decimalLatitude:32.8302; decimalLongitude:-17.1585; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:3 females
-
locationID: 24; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal - Levada dos Cedros; verbatimElevation:820; decimalLatitude:32.8259; decimalLongitude:-17.158; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:3 males, 4 females
-
locationID: 25; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:890; decimalLatitude:32.8236; decimalLongitude:-17.156; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 25; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:890; decimalLatitude:32.8236; decimalLongitude:-17.156; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 5 females
-
locationID: 26; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:889; decimalLatitude:32.8226; decimalLongitude:-17.1539; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 2 females
-
locationID: 27; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1023; decimalLatitude:32.8182; decimalLongitude:-17.1521; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 27; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1023; decimalLatitude:32.8182; decimalLongitude:-17.1521; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 28; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1134; decimalLatitude:32.8062; decimalLongitude:-17.1409; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 3 females
-
locationID: 28; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1134; decimalLatitude:32.8062; decimalLongitude:-17.1409; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 females
-
locationID: 29; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Ginjas; verbatimElevation:869; decimalLatitude:32.7758; decimalLongitude:-17.0534; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 29; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Ginjas; verbatimElevation:869; decimalLatitude:32.7758; decimalLongitude:-17.0534; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 33; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Rabaçal; verbatimElevation:930; decimalLatitude:32.7647; decimalLongitude:-17.1341; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 33; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Rabaçal; verbatimElevation:930; decimalLatitude:32.7647; decimalLongitude:-17.1341; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:4 females
-
locationID: 35; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Casa do Elias; verbatimElevation:814; decimalLatitude:32.8268; decimalLongitude:-17.1883; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 1 female
Dipoenata cf. longitarsis
-
locationID: 7; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Tanque; verbatimElevation:845; decimalLatitude:32.7425; decimalLongitude:-16.9168; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
Madeira island (Fig.
Enoplognatha sattleri
-
locationID: 11; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1211; decimalLatitude:32.7733; decimalLongitude:-16.9081; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 21; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Levada; verbatimElevation:568; decimalLatitude:32.8047; decimalLongitude:-16.9346; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 25; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:890; decimalLatitude:32.8236; decimalLongitude:-17.156; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
Episinus maderianus
-
locationID: 1; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:500; decimalLatitude:32.7493; decimalLongitude:-16.8114; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 1 female
-
locationID: 4; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:630; decimalLatitude:32.7415; decimalLongitude:-16.9161; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 1 female
-
locationID: 7; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Tanque; verbatimElevation:845; decimalLatitude:32.7425; decimalLongitude:-16.9168; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 9; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Queimadas; verbatimElevation:841; decimalLatitude:32.7873; decimalLongitude:-16.9047; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 10; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico das Pedras; verbatimElevation:883; decimalLatitude:32.7841; decimalLongitude:-16.9055; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 males
-
locationID: 12; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1103; decimalLatitude:32.7762; decimalLongitude:-16.9022; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 13; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Viveiro; verbatimElevation:906; decimalLatitude:32.7354; decimalLongitude:-16.8864; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 males
-
locationID: 14; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Cottages; verbatimElevation:994; decimalLatitude:32.7319; decimalLongitude:-16.8861; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 17; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:519; decimalLatitude:32.7933; decimalLongitude:-17.1122; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 21; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Levada; verbatimElevation:568; decimalLatitude:32.8047; decimalLongitude:-16.9346; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 4 females
-
locationID: 22; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Ribeiro; verbatimElevation:560; decimalLatitude:32.7985; decimalLongitude:-16.936; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 1 female
-
locationID: 25; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:890; decimalLatitude:32.8236; decimalLongitude:-17.156; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 27; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1023; decimalLatitude:32.8182; decimalLongitude:-17.1521; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 1 female
-
locationID: 28; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1134; decimalLatitude:32.8062; decimalLongitude:-17.1409; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 33; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Rabaçal; verbatimElevation:930; decimalLatitude:32.7647; decimalLongitude:-17.1341; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 34; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Ponta do Sol; locality:Risco; verbatimElevation:1048; decimalLatitude:32.7608; decimalLongitude:-17.1256; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 35; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Casa do Elias; verbatimElevation:814; decimalLatitude:32.8268; decimalLongitude:-17.1883; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 male, 1 female
Madeira island (Fig.
Previous records of E. maderianus were made by Schenkel (
Paidiscura orotavensis
-
locationID: 11; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1211; decimalLatitude:32.7733; decimalLongitude:-16.9081; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
Rugathodes madeirensis
-
locationID: 3; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Pte. Roquete; verbatimElevation:1074; decimalLatitude:32.7391; decimalLongitude:-16.9156; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 4; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:630; decimalLatitude:32.7415; decimalLongitude:-16.9161; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 8; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Til Gigante; verbatimElevation:841; decimalLatitude:32.7457; decimalLongitude:-16.915; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 9; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Queimadas; verbatimElevation:841; decimalLatitude:32.7873; decimalLongitude:-16.9047; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 10; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico das Pedras; verbatimElevation:883; decimalLatitude:32.7841; decimalLongitude:-16.9055; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 11; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1211; decimalLatitude:32.7733; decimalLongitude:-16.9081; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 12; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1103; decimalLatitude:32.7762; decimalLongitude:-16.9022; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 12; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1103; decimalLatitude:32.7762; decimalLongitude:-16.9022; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 13; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Viveiro; verbatimElevation:906; decimalLatitude:32.7354; decimalLongitude:-16.8864; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 17; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:519; decimalLatitude:32.7933; decimalLongitude:-17.1122; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 22; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Ribeiro; verbatimElevation:560; decimalLatitude:32.7985; decimalLongitude:-16.936; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:3 females
-
locationID: 24; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal - Levada dos Cedros; verbatimElevation:820; decimalLatitude:32.8259; decimalLongitude:-17.158; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 28; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1134; decimalLatitude:32.8062; decimalLongitude:-17.1409; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:1 female
-
locationID: 29; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:São Vicente; locality:Ginjas; verbatimElevation:869; decimalLatitude:32.7758; decimalLongitude:-17.0534; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
Madeira island (Fig.
Steatoda nobilis
-
locationID: 4; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:630; decimalLatitude:32.7415; decimalLongitude:-16.9161; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
Theridion melanurum
-
locationID: 4; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:630; decimalLatitude:32.7415; decimalLongitude:-16.9161; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 7; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Tanque; verbatimElevation:845; decimalLatitude:32.7425; decimalLongitude:-16.9168; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 9; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Queimadas; verbatimElevation:841; decimalLatitude:32.7873; decimalLongitude:-16.9047; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 11; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Achada do Teixeira; verbatimElevation:1211; decimalLatitude:32.7733; decimalLongitude:-16.9081; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 3 females
-
locationID: 14; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Cottages; verbatimElevation:994; decimalLatitude:32.7319; decimalLongitude:-16.8861; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 15; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1533; decimalLatitude:32.7231; decimalLongitude:-16.9109; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 26; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:889; decimalLatitude:32.8226; decimalLongitude:-17.1539; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:2 females
-
locationID: 27; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:1023; decimalLatitude:32.8182; decimalLongitude:-17.1521; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
Theridion n. sp.
-
locationID: 1; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:500; decimalLatitude:32.7493; decimalLongitude:-16.8114; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 1 female
-
locationID: 2; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Machico; locality:Funduras; verbatimElevation:552; decimalLatitude:32.754; decimalLongitude:-16.8099; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 4; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Fajã da Nogueira - Mtdo. do Leacoque; verbatimElevation:630; decimalLatitude:32.7415; decimalLongitude:-16.9161; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 13; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Viveiro; verbatimElevation:906; decimalLatitude:32.7354; decimalLongitude:-16.8864; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 14; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Frio - Cottages; verbatimElevation:994; decimalLatitude:32.7319; decimalLongitude:-16.8861; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 18; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Chão da Ribeira; verbatimElevation:491; decimalLatitude:32.7957; decimalLongitude:-17.1117; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 21; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Ribeiro Bonito - Levada; verbatimElevation:568; decimalLatitude:32.8047; decimalLongitude:-16.9346; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 female
-
locationID: 24; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal - Levada dos Cedros; verbatimElevation:820; decimalLatitude:32.8259; decimalLongitude:-17.158; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 33; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Rabaçal; verbatimElevation:930; decimalLatitude:32.7647; decimalLongitude:-17.1341; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male, 1 female
-
locationID: 34; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Ponta do Sol; locality:Risco; verbatimElevation:1048; decimalLatitude:32.7608; decimalLongitude:-17.1256; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
Madeira island (Fig.
This is a new species to science. It seems to be restricted to Laurisilva and is now being formally described (Van Keer, in prep.).
Misumena spinifera
-
locationID: 25; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Porto Moniz; locality:Fanal; verbatimElevation:890; decimalLatitude:32.8236; decimalLongitude:-17.156; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
Xysticus nubilus
-
locationID: 15; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1533; decimalLatitude:32.7231; decimalLongitude:-16.9109; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:2 females
Zodarion styliferum
-
locationID: 15; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Santana; locality:Pico do Areeiro; verbatimElevation:1533; decimalLatitude:32.7231; decimalLongitude:-16.9109; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:201 males, 63 females
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Direct sampling; sex:1 male
-
locationID: 32; higherGeography:Macaronesia; continent:Europe; waterBody:Atlantic Ocean; islandGroup:Madeira archipelago; island:Madeira; country:Portugal; countryCode:PT; stateProvince:Madeira; county:Calheta; locality:Rabaças; verbatimElevation:993; decimalLatitude:32.7413; decimalLongitude:-17.0783; geodeticDatum:WGS84; samplingProtocol:Pitfall; sex:14 males, 7 females
Discussion
Fifty two spider species and 3145 specimens are accounted in this study. Out of these, 17 are Madeira’s endemics species (SIE) mostly restricted to Laurisilva. Range for most endemic spider species was expanded by this study (Figs
Species richness considerations
The highest values of species richness were found in sites 18, 7, 11, 12, 25 and 33, which were also among those with the largest number of endemics (Fig.
Species richness and abundance per study site. Sampling methods used, number of endemic (SIE), native and introduced species are presented for each site. Site codes as in Table
| Site | Species richness (S) | Abundance (N) | Sampling methods | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Code | SIE | Native | Invasive | total | SIE | Native | Invasive | total | |
| 1 | 4 | 2 | 2 | 8 | 7 | 3 | 24 | 34 | Pitfall & direct sampling |
| 2 | 3 | 3 | 4 | 10 | 4 | 12 | 63 | 79 | Pitfall & direct sampling |
| 3 | 1 | 1 | 2 | 2 | 1 | 1 | 2 | 4 | Pitfall |
| 4 | 5 | 4 | 3 | 12 | 13 | 8 | 38 | 59 | Pitfall & direct sampling |
| 5 | 0 | 1 | 1 | 2 | 0 | 3 | 1 | 4 | Pitfall |
| 6 | 5 | 6 | 2 | 13 | 12 | 7 | 36 | 55 | Pitfall & direct sampling |
| 7 | 7 | 4 | 3 | 13 | 10 | 6 | 21 | 37 | Pitfall & direct sampling |
| 8 | 2 | 2 | 2 | 6 | 2 | 2 | 19 | 23 | Pitfall & direct sampling |
| 9 | 3 | 1 | 3 | 7 | 3 | 3 | 10 | 16 | Pitfall & direct sampling |
| 10 | 3 | 1 | 2 | 6 | 4 | 4 | 9 | 17 | Pitfall & direct sampling |
| 11 | 5 | 7 | 2 | 14 | 5 | 25 | 17 | 47 | Pitfall & direct sampling |
| 12 | 7 | 5 | 2 | 14 | 12 | 11 | 8 | 31 | Pitfall & direct sampling |
| 13 | 6 | 5 | 2 | 13 | 9 | 10 | 7 | 26 | Pitfall & direct sampling |
| 14 | 5 | 4 | 3 | 12 | 7 | 4 | 11 | 22 | Pitfall & direct sampling |
| 15 | 0 | 4 | 7 | 11 | 0 | 28 | 364 | 392 | Pitfall |
| 16 | 0 | 3 | 7 | 10 | 0 | 45 | 215 | 260 | Pitfall |
| 17 | 3 | 3 | 2 | 8 | 4 | 4 | 14 | 22 | Pitfall & direct sampling |
| 18 | 9 | 5 | 3 | 16 | 17 | 17 | 25 | 59 | Pitfall & direct sampling |
| 19 | 0 | 1 | 1 | 2 | 0 | 3 | 3 | 6 | Pitfall & direct sampling |
| 20 | 1 | 1 | 3 | 5 | 1 | 2 | 13 | 16 | Pitfall |
| 21 | 5 | 2 | 2 | 9 | 17 | 3 | 37 | 57 | Pitfall & direct sampling |
| 22 | 4 | 1 | 2 | 7 | 8 | 1 | 20 | 29 | Pitfall & direct sampling |
| 23 | 0 | 0 | 2 | 2 | 0 | 0 | 5 | 5 | Pitfall |
| 24 | 3 | 5 | 3 | 11 | 7 | 22 | 22 | 51 | Pitfall & direct sampling |
| 25 | 6 | 6 | 2 | 8 | 11 | 12 | 19 | 42 | Pitfall & direct sampling |
| 26 | 0 | 2 | 3 | 5 | 0 | 7 | 7 | 14 | Pitfall & direct sampling |
| 27 | 2 | 4 | 3 | 9 | 3 | 7 | 9 | 19 | Pitfall & direct sampling |
| 28 | 5 | 4 | 3 | 12 | 5 | 9 | 15 | 29 | Pitfall & direct sampling |
| 29 | 4 | 2 | 2 | 8 | 6 | 5 | 11 | 22 | Pitfall & direct sampling |
| 30 | 1 | 1 | 1 | 3 | 1 | 2 | 2 | 5 | Pitfall |
| 31 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | Pitfall |
| 32 | 0 | 6 | 6 | 12 | 0 | 11 | 49 | 60 | Pitfall |
| 33 | 7 | 5 | 2 | 14 | 11 | 7 | 17 | 35 | Pitfall & direct sampling |
| 34 | 7 | 3 | 1 | 11 | 9 | 3 | 8 | 20 | Pitfall & direct sampling |
| 35 | 1 | 0 | 2 | 3 | 2 | 0 | 5 | 7 | Pitfall |
| 36 | 1 | 1 | 1 | 3 | 1 | 2 | 12 | 15 | Pitfall |
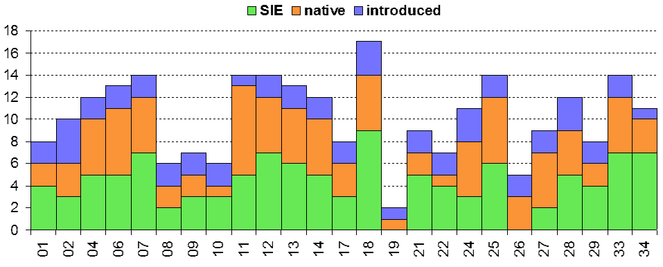
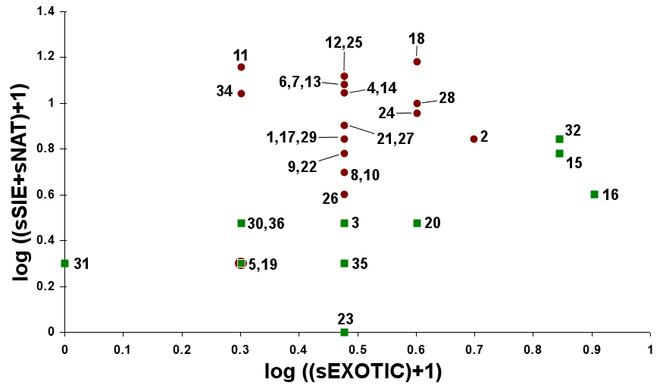
This study reveals a high number of endemic spider species in some locations, such as Chão da Ribeira (sites 17 and 18), Rabaçal (site 33), Risco (site 34), Achada do Teixeira (sites 11 and 12), Ribeiro Frio (sites 13 and 14), Fajã da Nogueira (sites 3 to 8) and Fanal (sites 23 to 28) that should therefore be the focus of further conservation biology of Madeiran spiders and their associated habitats. Those sites are the only where Centromerus variegatus, Ceratinopsis infuscata, Dipoenata longitarsis, Frontinellina dearmata, Frontiphantes fulgurenotatus and Lepthyphantes mauli are known to occur. However, the occurrence of these araneoid spiders in other locations within Madeira island is expected since they have a potentially high dispersal ability.
This is the first time that a large set of standardized samples of spiders from Madeira island is reported. The distribution of several Madeiran endemic species was greatly increased, particularly Macarophaeus cultior, Hahnia insulana, Turinyphia maderiana, Trogloneta madeirensis, Rugathodes madeirensis and Episinus maderianus. Other endemics were recorded in fewer sites, but in most cases such species are still associated with Laurisilva. In some instances, the historical distribution of endemic spider species includes areas no longer covered by Laurisilva. An example is a record of Centromerus variegatus from Santo da Serra (
The need to apply a combination of sampling techniques in a standardized design in order to assess species composition and structure of ecological communities has been strongly emphasized in the last few years by different authors (
In complex habitats like Laurisilva, the prospection of microhabitats such as fallen logs and trees, tree holes, under tree bark and epiphytes allows the finding of species with highly specialized niches. Furthermore, several spider species seldom move or do not move at all at ground level, making direct sampling the most suitable detection technique. From the 52 spider species reported, 16 were collected exclusively using pitfall traps while 22 other species were only found by direct sampling. On the other hand, direct sampling proved to be an effective method to sample Madeiran endemics since most of the sampled endemic spider species were only collected using this technique (see also Fig.
The original descriptions of some Madeiran endemics do not allow inequivocal identification since they were based on very few specimens or on specimens of a single gender. Thus, taxonomic revisions of several genera or families (i.e. Dysdera Latreille, 1804, Hahnia C.L. Koch, 1841, or the Linyphiidae) are mandatory for a better understanding of Madeiran spider biodiversity. Besides the merits of the present study in improving the knowledge on the distribution of spider species in Madeira island, it also allowed the collection of a number of specimens that will be the aim of future taxonomic studies, such as species redescriptions and formal descriptions of new species to science.
Madeira Laurisilva presents a unique spider biodiversity and efforts should be made towards improving knowledge on the abundance and distribution of endemic species as well as to develop a strategy for their conservation and monitoring.
Acknowledgements
We acknowledge Madeira Natural Park, particularly Herculano Fernandes and Martinho Gomes, for the valuable support during fieldwork. Financial support was provided by Fundação para a Ciência e Tecnologia (FCT, Portugal) through projects PDCT/BIA-BDE/59202/2004 and PTDC/BIABEC/99138/2008 (http://www.fct.pt/). CR was supported by grant SFRH/BPD/91357/2012, and IR by grants SFRH/BPD/29578/2006 (FCT, Portugal) and M3.1.7/F/030/2011 (FRCT, Açores, Portugal).
References
- Descriptions of newly discovered spiders captured by James Yate Johnson Esq., in the island of Madeira.Annals and Magazine of natural History4:255‑267.
- Descriptions of newly-discovered spiders from the island of Madeira.Annals and Magazine of Natural History9:370‑382.
- A list of the terrestrial fungi, flora and fauna of Madeira and Selvagens archipelagos.Direcção Regional do Ambiente da Madeira and Universidade dos Açores,Funchal and Angra do Heroísmo,440pp.
- Ranking protected areas in the Azores using standardized sampling of soil epigean arthropods.Biodiversity and Conservation14:2029‑2060. https://doi.org/10.1007/s10531-004-4283-y
- Spider biodiversity patterns and their conservation in the Azorean archipelago, with descriptions of new species.Systematics and Biodiversity6(2):249‑282. https://doi.org/10.1017/S1477200008002648
- Invasibility and species richness of island endemic arthropods: a general model of endemic vs. exotic species.Journal of Biogeography33(1):169‑187. https://doi.org/10.1111/j.1365-2699.2005.01324.x
- Beitrag zur Kenntnis der Arachniden-Fauna von Madeira und den Canarischen Inseln.Abhandlungen naturwissenschaftlichen Vereins Hamburg13:1‑13.
- Spiders collected by the Shackleton-Rowett Expedition in the island of Madeira.Annals and Magazine of Natural History15:331‑334.
- Standardization and optimization of arthropod inventories — the case of Iberian spiders.Biodiversity and Conservation18(14):3949‑3962. https://doi.org/10.1007/s10531-009-9690-7
- Cardoso P, Crespo LC (2008) Araneae. In: Borges PA, Abreu C, Aguiar AM, Carvalho P, Jardim R, Melo I, Oliveira P, Sérgio C, Serrano AR, Vieira P (Eds) A list of the terrestrial fungi, flora and fauna of Madeira and Selvagens archipelagos.Direcção Regional do Ambiente da Madeira and Universidade dos Açores,Funchal and Angra do Heroísmo,440pp.
- Cardoso P, Wunderlich J, Borges PA (2010) Araneae. In: Borges PA, Costa A, Cunha R, Gabriel R, Gonçalves V, Martins AF, Melo I, Parente M, Raposeiro P, Rodrigues P, Santos RS, Silva L, Vieira P, Vieira V (Eds) A list of the terrestrial and marine biota of the Azores.Princípia,Cascais,432pp.
- The seven impediments in invertebrate conservation and how to overcome them.Biological Conservation144(11):2647‑2655. https://doi.org/10.1016/j.biocon.2011.07.024
- Ad-Hoc vs. Standardized and Optimized Arthropod Diversity Sampling.Diversity1(1):36‑51. https://doi.org/10.3390/d1010036
- Rapid biodiversity assessment, faunistics and description of a new spider species (Araneae) from Desertas Islands and Madeira (Portugal).Revista Ibérica de Aracnología23:11‑23.
- Les araignées de l’archipel de Madère (Mission du Professeur Vandel).Publicações do Instituto de Zoologia Dr. Augusto Nobre79:1‑118.
- Spiders from the Madeira and Salvage Islands.Boletim do Museu Municipal do Funchal17:29‑48.
- Spiders from Azores and Madeira.Boletim do Museu Municipal do Funchal18:68‑102.
- IUCN evaluation of nominations of natural and mixed properties to the World Heritage List.The 23rd ordinary session of the World Heritage Commitee,Paris, France,5-10 July 1999.
- Description of a new species of Lycosa living in the island of Madeira; with some remarks on the Lycosa tarentuloides maderiana Walckenaer.Annals and Magazine of Natural History12:152‑155.
- Arachnoidea opera Rev. Schmitz collecta in insulis Maderianis et Selvages dictis.Rozprawa i sprawodzanie wydziat matematyczno-pzyrodniczy Academia umiejetnosi Cracov36:319‑461.
- Araneae nonnullae in insulus Maderianis collectae a Rev. E. Schmitz.Bulletin de l’Academie des Sciences de Cracovie1905:440‑460.
- A review of the genus Pellenes in the fauna of Central Asia and the Caucasus (Araneae, Salticidae).Journal of Natural History33:89‑148. https://doi.org/10.1080/002229399300489
- Lomolino MV (2004) Conservation Biogeography. In: Lomolino MV, Heaney LR (Eds) Frontiers of biogeography: new directions in the geography of Nature.Sinauer Associates, Inc.,Sunderland, MA.
- Descriptions of two species of Araneidae, natives of Madeira.Zoological Journal5:320‑323.
- Macías N (2009) Araneae. In: Arechavaleta M, Rodríguez S, Zurita N, García A (Eds) Lista de especies silvestres de Canarias. Hongos, plantas y animales terrestres.Gobierno de Canarias.,Santa Cruz de Tenerife.
- Using taxonomically unbiased criteria to prioritize resource allocation for oceanic island species conservation.Biodiversity and Conservation19(6):1659‑1682. https://doi.org/10.1007/s10531-010-9795-z
- Top 100 – As cem espécies ameaçadas prioritárias em termos de gestão na região europeia biogeográfica da Macaronésia.Consejería de Medio Ambiente y Ordenación Territorial, Gobierno de Canarias,500pp.
- Biodiversity hotspots for conservation priorities.Nature203:853‑858. https://doi.org/10.1038/35002501
- Laurisilva da Madeira – caracterização quantitativa e qualitativa.Parque Natural da Madeira, Grafimadeira,Funchal,192pp.
- The world spider catalog.14.0.American Museum of Natural History. URL: http://research.amnh.org/entomology/spiders/catalog/index.html
- Die Arthropodenfauna von Madeira nach den Ergebnissen der Reise von Prof. Dr. O. Lundblad, 1935.Arkiv för Zoologi30:1‑42.
- Arachnidos da Madeira.Annaes des Sciencias Naturaes2:197‑199.
- IFRAM1 – 1º Inventário Florestal da Região Autónoma da Madeira.Direcção Regional de Florestas,Funchal,120pp.
- Spiders from Madeira.Annals and Magazine of Natural History10:216‑228.
- Die Spinnen der Kanarischen Inseln und Madeiras: Adaptive Radiation, Biogeographie, Revisionen und Neubeschreibungen.Triops Verlag,Langen, West Germany,435pp.
- Die Spinnen-Fauna der Makaronesischen Inseln.Beiträge zur Araneologie1:1‑619.
- Zu Ökologie, Biogeographie, Evolution und Taxonomie einiger Spinnen der Makaronesischen Inseln (Arachnida: Araneae).Beiträge zur Araneologie4:385‑439.
- Extant and fossil spiders (Araneae).Beiträge zur Araneologie6:1‑640.
Supplementary material
Data from where Figures 5 and 6 were built.