|
Biodiversity Data Journal :
Taxonomic Paper
|
|
Corresponding author: Jukka Salmela (jukka.e.salmela@gmail.com)
Academic editor: Vladimir Blagoderov
Received: 10 Jan 2017 | Accepted: 01 Mar 2017 | Published: 06 Mar 2017
© 2017 Jukka Salmela, Levente-Péter Kolcsár
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Salmela J, Kolcsár L (2017) New and poorly known Palaearctic fungus gnats (Diptera, Sciaroidea). Biodiversity Data Journal 5: e11760. https://doi.org/10.3897/BDJ.5.e11760
|
|
Abstract
Background
Fungus gnats (Sciaroidea) are a globally species rich group of lower Diptera. In Europe, Fennoscandian peninsula in particular holds a notable diversity, ca. 1000 species, of which 10 % are still unnamed. Fungus gnats are predominantly terrestrial insects, but some species dwell in wetland habitats.
New information
Eight new fungus gnat species, belonging to the families Keroplatidae (Orfelia boreoalpina Salmela sp.n.) and Mycetophilidae (Sciophila holopaineni Salmela sp.n., S. curvata Salmela sp.n., Boletina sasakawai Salmela & Kolcsár sp.n., B. norokorpii Salmela & Kolcsár sp.n., Phronia sompio Salmela sp.n., P. reducta Salmela sp.n., P. prolongata Salmela sp.n.), are described. Four of the species are known from Fennoscandia only whilst two are supposed to have boreo-alpine disjunct ranges, i.e. having populations in Fennoscandia and the Central European Alps. One of the species probably has a boreal range (Finnish Lapland and Central Siberia). Type material of Boletina curta Sasakawa & Kimura from Japan was found to consist of two species, and a further species close to these taxa is described from Finland. Phronia elegantula Hackman is redescribed and reported for the first time from Norway. DNA barcodes are provided for the first time for five species.
Keywords
boreal zone, boreo-alpine species, DNA barcoding, biodiversity, taxonomy
Introduction
Sciaroidea are lower Diptera traditionally classified to the infraorder Nematocera, thread-horned flies (
Fungus gnats are a highly diverse group of flies, having over 5000 known species globally (
DNA barcoding has become a standard procedure in biodiversity surveys and taxonomic studies (e.g.
The new species described here belong to the genera Orfelia Costa, Sciophila Meigen, Boletina Staeger and Phronia Winnertz. Orfelia is a keroplatid genus with 36 Holarctic species, of which 25 are known from the Palaearctic region (
Materials and methods
Most of the specimens studied were collected from Finland, mainly by using Malaise traps. Ethylene glycol was first used in the traps as a preservative and later the material was stored in 70 % ethanol. The morphological terminology used here follows (
Images of male hypopygia were taken using an Olympus SZ61 stereomicroscope equipped with a Canon 650D camera and a LM Digital SLR Adapter. Habitus photos of the new Orfelia species were taken by using Olympus E520 digital camera, attached to an Olympus SZX16 stereomicroscope. Digital photos were captured using the programmes Deep Focus 3.1 and Quick PHOTO CAMERA 2.3. Extended depth of field photos were reconstructed using the software Combine ZP and were finalized with the use of Adobe Photoshop CS4. The maps were drawn by using SimppleMappr program (http://www.simplemappr.net/).
A 658 bp fragment of mitochondrial protein-encoding cytochrome c oxidase subunit I (COI) was sequenced from a total of 10 Sciaroidea specimens. Legs or 2–3 abdominal segments of the specimens were placed in 96% ethanol in a 96-well lysis microplate and dispatched to the Canadian Centre for DNA Barcoding, Biodiversity Institute of Ontario where DNA was extracted and sequenced using standard protocols and primers (
Barcodes of the Finnish specimens (all 658 bp in length, with no unambiguous bases) were submitted to the BOLD (
Taxon treatments
Orfelia boreoalpina , sp. n.
-
phylum: Arthropoda; class:Insecta; order:Diptera; country:Finland; stateProvince:Lapponia kemensis pars orientalis; municipality:Savukoski; locality:Törmäoja Conservation Area; decimalLatitude:67.823; decimalLongitude:29.439; eventDate:2014-08-07; individualCount:1; sex:M; lifeStage:adult; catalogNumber:DIPT-JS-2014-0233; recordedBy:M. Mäkilä; identifiedBy:Jukka E. Salmela; institutionCode:ZMUT
-
country: Germany; stateProvince:Bavaria; locality:Nationalpark Bayerischer Wald, 11.3 km N of Grafenau; decimalLatitude:48.9509; decimalLongitude:13.422; eventDate:2012-09-13/22; individualCount:1; sex:female; catalogNumber:BIOUG08366-D12; recordNumber:bayw.17; recordedBy:G. Sellmayer; institutionCode:ZSM
Description
Male. Head bicolored, vertex with a triangular dark area, laterally yellowish brown (Fig.



Scutum yellowish with three longitudinal brown stripes; median stripe consisting of two stripes that are largely merged, a narrow anterior gap between the stripes is present (Fig.
Wings yellowish, with a faint subapical dark band extending from C to M2. Veins dark brown except bm–cu and bRs that are lighter. Veins R1 and bCuA with dorsal setae, R5 setose both ventrally and dorsally. Sc ending in C before bRs. R4 very short, about 0.12 times longer than apical portion of R5. Wing length 4.1 mm.
Coxae yellowish brown - brown, bearing short dark setae, legs yellowish. Ratio of femur to tibia for fore, mid and hind legs: 0.79, 0.68, 0.63. Ratio of tibia to basitarsus for fore, mid and hind legs: 1.67, 1.0, 1.0. Anterior spur of mid-tarsus about 0.5 times longer than posterior spur.
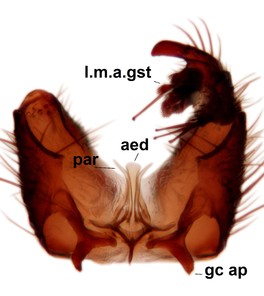
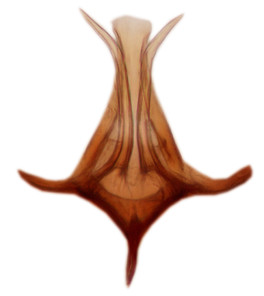
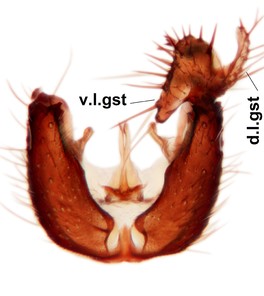
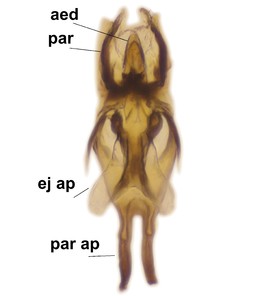
Abdominal tergites and sternites brown, bearing dark setae. Hypopygium brown. 9th tergite widest medially, apex rounded. Gonocoxites dorsally with an outgrowth, bearing a few long apical setae and having a mesial protrusion (Fig.
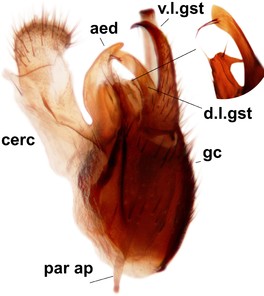
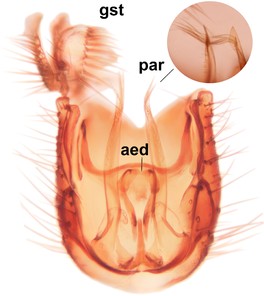
Orfelia boreoalpina Salmela sp.n., holotype male, hypopygium.
b: Hypopygium, ventral view.
c: Parameres (par), ventral view.
d: Aedeagus, ventral view.
Female.The paratype female is lacking all legs except right fore leg and right hind femur. The specimen is slightly paler than the holotype male. The specimen may be somewhat teneral or it has bleached in the Malaise trap or later in the ethanol. Otherwise the specimen is very similar to the holotype. Antennal flagellomeres, except first and last, are wider than long (length:width ratio of 4th segment is 0.78). Cerci short, apically truncated, gonocoxite 8 short and rounded. Wing length 3.9 mm.
Diagnosis
The new species is characterised by the short and dark antennae, a yellow scutum with contrasting scutellar stripes, brown pleural sclerites of the thorax, brown, unicolorous abdomen and a short R4 vein. The dorsal lobe of gonostylus is strongly curved. The ventral lobe of gonostylus has only two black apical setae, while O. nigricornis (Fabricius) and O. subnigricornis Zaitzev & Menzel have a bunch of setae.
Etymology
The name of the new species refers to its putative boreo-alpine, disjunct range in Europe. The name is a noun in apposition.
Distribution
The new species has been observed from eastern Finnish Lapland, the north boreal ecoregion, and from Germany, Bavaria (see
Ecology
The Finnish sampling site was a herb-rich meadow, harbouring vascular plants such as Bistorta vivipara and Trollius europaeus, and is probably flooded during snowmelt in spring. The meadow is surrounded by pine (Pinus sylvestris) dominated boreal forest. Bavarian site is a conifer-dominated mountain forest (
Taxon discussion
The new species is rather distant to all other Holarctic species, but it may be closest to O. nigricornis and O. subnigricornis (see below). If using the key provided by
DNA barcoding
Holotype male: BOLD Sample ID: DIPT-JS-2014-0233. BOLD Process ID: SCFI064-15. GenBank accession number: KY062990.
AACATTATATTTTATTTTAGGGACATGGTCAGGAATACTAGGAACATCAATAAGAATTTTAATTCGAGCAGAATTAGGATATCCGGGAGCATTAATTGGAAACGACCAAATTTATAATGTTGTAGTCACAGCTCATGCTTTTGTAATAATTTTTTTTATAGTTATACCTACTATAATTGGAGGTTTCGGAAATTGATTAGTACCTTTAATATTAGGGGCCCCAGATATGGCTTTTCCTCGAATAAATAACATAAGATTTTGACTTCTCCCTCCTTCACTTTCTTTACTATTAATAAGAAGAATAGTAGAAAGTGGTTCTGGAACAGGATGAACTGTATATCCTCCCCTATCTTCTACTTTATCTCATTCTGGTAGATCAGTTGACTTAACTATTTTTTCTCTTCATTTAGCAGGAATTTCTTCAATTCTTGGGGCAGTCAATTTTATTACTACAATTATCAACATACGATCACCTGGGATAAACATAGACATAATACCTTTATTTGTATGATCAGTTTTTATTACAGCCATTCTTCTTCTTTTATCATTACCTGTACTAGCGGGAGCAATTACAATACTTTTAACAGATCGTAATTTAAATACATCATTTTTTGATCCAGCAGGTGGGGGTGACCCAATTCTATATCAACATTTATTT
The DNA barcode of the paratype specimen is almost identical to the holotype, their similarity is 99.54 %. The type specimens belong to the same BIN (BOLD:ACJ7389) shared by no other members. The nearest specimens are rather distant: 97 closest sequences have similarity values between 88.25 and 86.35, being assigned to O. nemoralis (Meigen) (54 specimens), O. nigricornis (2), Keroplatidae (40) and Mycetophilidae (1). DNA barcode and associated data of the paratype is available from the BOLD Public data portal.
Sciophila holopaineni , sp. n.
-
country: Finland; stateProvince:Lapponia kemensis pars orientalis; verbatimLocality:Törmäoja Conservation Area, Hannu Ollin vaara; verbatimLatitude:67.843; verbatimLongitude:29.468; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2013-7-8/9-19; habitat:old-growth boreal forest, dominated by birch (Betula sp.); individualCount:1; sex:male; catalogNumber:DIPT-JS-2015-0075; recordedBy:J. Salmela; identifiedBy:J. Salmela; institutionCode:ZMUT
-
country: Russia; stateProvince:Karelia; verbatimLocality:2 km NW of Syrovatka island; verbatimLatitude:65.528; verbatimLongitude:34.729; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2003-7-20/22; habitat:sea-shore meadow, close to a forest margin; individualCount:1; sex:male; recordedBy:A. Polevoi; identifiedBy:J. Salmela; institutionCode:ZIN
Description
Male. Head black. Ocelli arranged in a row, on the posterior part of vertex; ratio of distance of lateral ocellus from median ocellus: distance of lateral ocelli from eye = 0.52. Vertex, anterior part of face and clypeus covered by dark setae. Eyes pubescent. Palpi infuscated, with dark setae. Length ratio of palpal segments 3–5: 3:4=0.94, 4:5=0.43. Penultimate segment 3.4 times as long as wide, last segment 9.3 times as long as wide. Antennae 16-segmented (scape, pedicel and 14 flagellomeres), black. Scape:pedicel length ratio 1.30; scape with a rounded, a bit depressed sensory field in its lateral base, having 7 minute setae. Flagellomeres cylindrical, length:width ratio of 1st flagellomere 1.51, 4th flagellomere 1.76 and apical flagellomere 3.13. Flagellomeres covered by dense light setosity, setae slightly curved, their length shorter than width of respective flagellomere; polygon-like (reticulate) pattern present, especially so in apical flagellomeres.
Thorax black. Scutum covered by pale setae. Anepimeron bare, other sclerites setose. Scutellum with eight setae in a curved row. Halteres light brown with pale setae; apical part of stem and base of knob infuscated.
Wings hyaline, lamina covered by both macro and microtrichia. Base of Rs, R4 and r-m bare, other veins setose, veins light brown to dark brown. C exceeding tip of R5 25 % of the distance between R5 and M1. Sc2 situated between base of Rs and R4. Furcation point of median fork at the level of bRs. M1+M2 very short. Length ratio of M1+2:r-m = 0.53. Wing length 3.2 mm.
Fore coxae light brown, mid and hind coxae dark brown, with pale setae, trochanters dark-brown. Legs yellowish brown, femora basoventrally darkened; apices of mid and hind coxae infuscated, the latter more clearly so. Setae on femora mostly dark, tibial and tarsal setae dark. Length ratio of femur to tibia for fore, mid and hind legs: 0.93, 1.03, 0.89. Length ratio of tibia to basitarsus for fore, mid and hind legs: 1.36, 1.57, 1.89. Anteroapical depressed area of the fore tibia with two rows of pale setae, proximal row curved with ca. 17 setae and distal row almost straight with ca. 20 setae. Ratio of apical width of tibia:length of longest tibial spur for fore, mid and hind legs: 0.52, 0.33, 0.33.
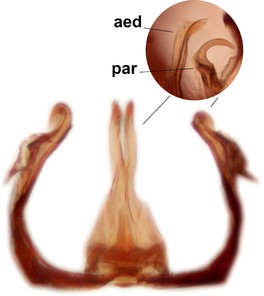
Abdominal tergites and sternites dark brown - almost black, covered by dark setae. Distal margin of 9th tergite rounded (Fig.


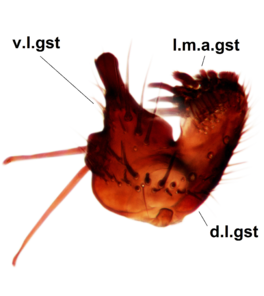
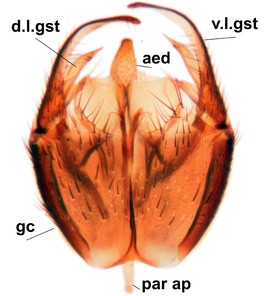
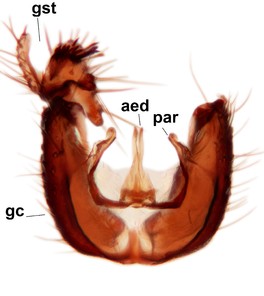
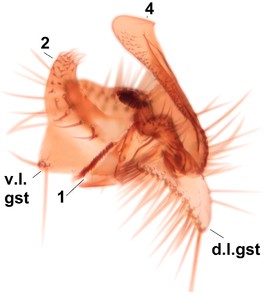
Sciophila holopaineni Salmela sp.n., holotype male, hypopygium.
b: Hypopygium, dorsal view. gc=gonocoxite, gst=gonostylus, l.m.a.gst=large median appendage of gonostylus with comb-like megasetae, par=parameres, aed=aedeagus, gc ap=gonocoxal apodeme.
c: Hypopygium, ventral view. v.l.gst=ventral lobe of gst.
d: Gonostylus, outer lateral view. d.l.gst=dorsal lobe of gonostylus.
e: Gonostylus, inner lateral view. s.m.a.gst=small median appendage of gst.
f: Aedeagus and parameres.
Diagnosis
This is a very dark species with the head, antennae, thorax, and abdomen black or dark brown. The 9th tergite is apically rounded. The ventral lobe of the gonostylus has a prominent apical outgrowth. The aedeagus is about as long as the parameres with the apex truncated. The parameres are rather thin with their apices contorted.
Etymology
The new species is named after Mr. Tuomas Holopainen, the founder, songwriter and keyboardist of a Finnish metal band, "Nightwish". The name is a genitive.
Distribution
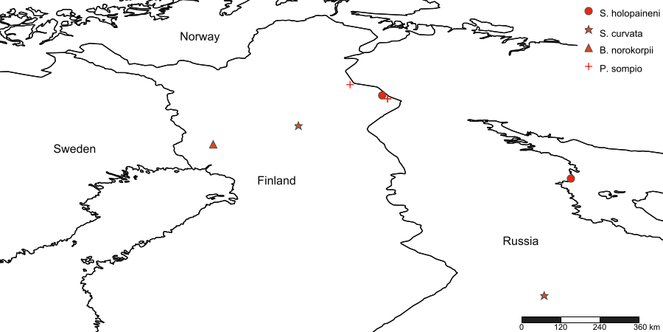
The new species is so far known only from eastern Finnish Lapland, the north boreal ecoregion (Fig.
Ecology
The type locality in Törmäoja Conservation Area was a sloping birch forest in a river canyon, close to a spring brook.
Taxon discussion
The new species seems to be rather distant from the known Holarctic species of Sciophila. The number of large setae on the small median appendage of gonostylus is varying, it may be two or three, thus making the use of Zaitzev's (
We were not able to find any notes in the literature on the presence of a sensory field at the base of scape among Sciophila. JS checked a few specimens in his collection (JES), and the character was present in S. buxtoni Freeman, S. curvata sp.n., Leptomorphus forcipatus Landrock, Polylepta borealis Lundström and Allocotocera pulchella (Curtis), but it was absent among Anaclileia dziedzickii (Landrock). In Acnemia trifida Zaitzev there was an ventroapical sensory field at the scape, with hyaline cover. It is possible, that this trait is symplesiomorphic (an ancestral character or trait state shared by two or more taxa) amongst Sciophilinae and is lost in some genera.
Sciophila curvata , sp. n.
-
country: Finland; stateProvince:Ostrobothnia borealis pars borealis; verbatimLocality:Kemijärvi, Pyhä-Luosto National Park, Karhunotko; verbatimLatitude:67.001; verbatimLongitude:27.133; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2014-6-10/7-11; habitat:old-growth boreal forest with an intermittent brook; individualCount:1; sex:male; catalogNumber:DIPT-JS-2015-0252; recordedBy:J. Salmela; identifiedBy:J. Salmela; institutionCode:ZMUT
-
country: Russia; stateProvince:Karelia; verbatimLocality:Kivach Nature Reserve; verbatimLatitude:62.272; verbatimLongitude:33.986; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:1990-8-13/9-11; habitat:Myrtillus pine forest; individualCount:1; sex:male; recordedBy:A. Polevoi; identifiedBy:J. Salmela; institutionCode:FRIP
Description
Male. Head dark, almost black. Ocelli arranged in a shallow triangle, approximately on the median part of vertex; ratio of distance of lateral ocellus from median ocellus: distance of lateral ocelli from eye = 0.59. Vertex covered by dark setae, face covered by small setae and clypeus by longer setae. Eyes pubescent. Palpi pale, covered by pale setae. Length ratio of palpal segments 3–5: 3:4=0.81, 4:5=0.52. Penultimate segment 4.3 times as long as wide, last segment 11.0 times as long as wide. Antennae 16-segmented (scape, pedicel and 14 flagellomeres), brown, first flagellomere light brown. Scape:pedicel length ratio 1.38. Scape with a slightly depressed sensory field in its base, having 5-6 minute setae. Flagellomeres cylindrical, length:width ratio of 1st flagellomere 1.54, 4th flagellomere 1.3 and apical flagellomere 2.90. Flagellomeres covered by dense light setosity, setae slightly curved, their length shorter than width of respective flagellomere. Polygonal (reticulate) pattern not present in basal and median flagellomeres, and either unclearly present or absent on the apical flagellomeres; apical flagellomeres of the holotype are slightly wrinkled.
Thorax dark brown. Scutum covered by pale setae. Anepimeron bare, other sclerites setose. Scutellum with eight setae in a curved row. Halteres light brown with pale setae.
Wings hyaline, both macro and microtrichia present on lamina. Base of Rs and R4 bare, other veins setose, veins brown to dark brown. C exceeding tip of R5 22 % of the distance between R5 and M1. Sc2 situated above R4. Furcation point of median fork slightly before the level of R4. Length ratio of M1+2:r-m = 0.71. Wing length 2.6 mm
Coxae yellow, with pale setae, trochanters infuscated. Legs yellow, femora ventrobasally darkened, setae on femora pale, tibial and tarsal setae darker. Length ratio of femur to tibia for fore, mid and hind legs: 0.98, 0.92, 0.83. Length ratio of tibia to basitarsus for fore, mid and hind legs: 1.81, 1.64, 2.20. Anteroapical depressed area of the fore tibia with ca. 16 pale setae in a row. Ratio of apical width of tibia:length of longest tibial spur for fore, mid and hind legs: 0.65, 0.27, 0.26.
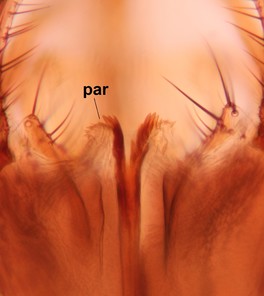
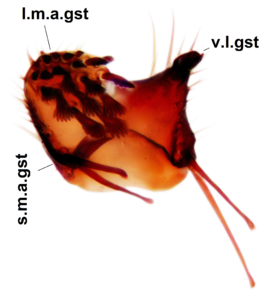
Abdominal tergites and sternites dark brown, covered by pale setae. 9th tergite triangular, apex pointed (Fig.


Sciophila curvata Salmela sp.n., holotype male, hypopygium.
b: Hypopygium, dorsal view. gc=gonocoxites, gst=gonostylus, aed=aedeagus, par=parameres.
c: Hypopygium, ventral view. v.l.gst=ventral lobe of gonostylus, d.l.gst=dorsal lobe of gonostylus.
d: Gonostylus, inner lateral view. l.m.gst=large median appendage of gonostylus, s.m.a.gst=small median appendage of gst.
e: Gonostylus, outer lateral view.
f: Aedeagus and parameres, dorsal view (insert shows lateral view on the apices of aedeagus and parameres).
Diagnosis
The new species is characterised by the presence of three setae on the small median lobe of the gonostylus, very narrow dorsal lobe of the gonostylus and strongly curved parameres. The new species is closest to S. californiensis Zaitzev; the 9th tergite of the latter species is medially constricted, in the former the outline of the 9th tergite is triangular.
Etymology
The name of the new species (curvata Latin, curved, an adjective) refers to the curved parameres of the male hypopygium.
Distribution
The type locality of the new species is from the Pyhä-Luosto National Park in central Finnish Lapland.
Ecology
The trapping site was a herb-rich bed of an intermittent brook, surrounded by an old-growth boreal forest.
Taxon discussion
The new species is most likely close to S. californiensis, because they share the following characters: the small median lobe of the gonostylus has three prominent setae and a narrow dorsal lobe. The new species, however, differs from S. californiensis by having a triangular 9th tergite (with a median constriction in S. californiensis) and having 18 comb-like megasetae (48 in S. californiensis). Although not mentioned in the description and improperly figured (
Boletina curta
Nomenclature
Sasakawa & Kimura, 1974: 60 (fig. 15a,b,c)
Zaitzev 1994: 209 (fig. 69,7)
-
country: Japan; stateProvince:Honsu; verbatimLocality:Otsu, Mt. Hiei; verbatimLatitude:35.06; verbatimLongitude:135.83; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; eventDate:1974-5-3; individualCount:1; sex:male; recordedBy:M. Sasakawa; identifiedBy:J. Salmela; institutionCode:OSAKA
Distribution
Boletina curta is a poorly known East Palaearctic species, hitherto recorded from Japan, Honshu (
Taxon discussion
Boletina curta was described from Honshu, the main island of Japan (
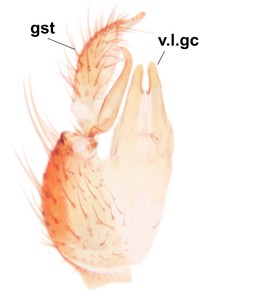
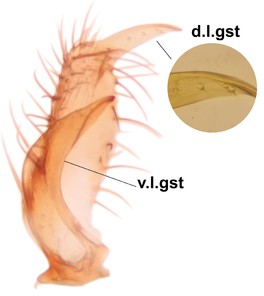
Boletina curta can be separated from the closely related B. sasakawai sp.n. and B. norokorpii sp.n. based on the following characters: 1) two stout setae present on the ventral lobe of the gonostylus (in the other taxa the ventral lobes of the gonostyli are bare, Fig.

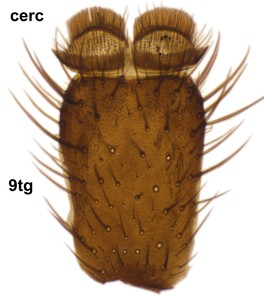
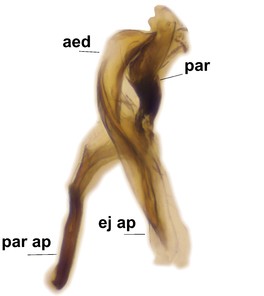
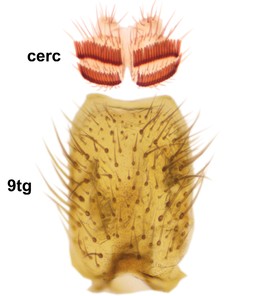
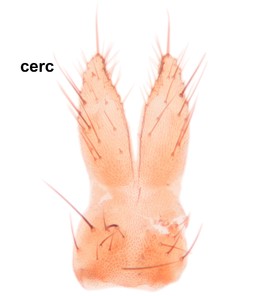
Boletina curta Sasakawa & Kimura, paratype male, hypopygium.
b: Cerci (cerc) and 9th tergite (9tg), dorsal view.
c: Gonostylus, lateral view. d.l.gst=dorsal lobe of gst, v.l.gst=ventral lobe of gst. Upper insert shows apex of d.l.gst and lower one two spines on the v.l.gst.
d: Aedeagal complex, dorsal view. aed=aedeagus, par=paramere, ej ap=ejaculatory apodeme, par ap=parameral apodeme.
e: Aedeagal complex, lateral view.



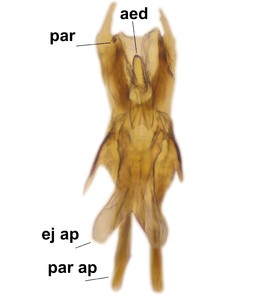

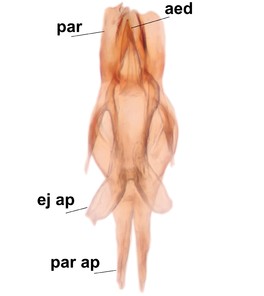
Boletina sasakawai Salmela & Kolcsár sp.n., holotype male, hypopygium.
b: Cerci (cerc) and 9th tergite (9tg), dorsal view.
c: Gonostlys, lateral view. d.l.gst=dorsal lobe of gonostylus, v.l.gst=ventral lobe of gonostylus.
d: Aedeagal complex, dorsal view. par ap=parameral apodemes, ej ap=ejaculatory apodemes, par=parameres, aed=aedeagus.
e: Aedeagal complex, lateral view.
Boletina norokorpii Salmela & Kolcsár sp.n., holotype male, hypopygium.
b: Cerci (cerc) and 9th tergite (9tg), dorsal view.
c: Gonostylus, lateral view. d.l.gst=dorsal lobe of gonostylus, v.l.gst=ventral lobe of gonostylus.
d: Aedeagal complex, dorsal view. aed=aedeagus, par=parameres, par ap=parameral apodemes, ej ap=ejaculatory apodemes.
e: Aedeagal complex, lateral view.
Some Boletina species, such as B. trivittata Staeger, occur in both early and late season (J. Salmela, pers.obs.). Hence, it might be possible in theory that B. sasakawai sp.n. is just a late summer/autumn morph of B. curta, likewise the butterfly species Araschnia levana (Linnaeus), that has two distinct colour morphs within a season (see e.g.
Boletina sasakawai , sp. n.
-
country: Japan; stateProvince:Honsu; verbatimLocality:Yoshino, Nara; verbatimLatitude:34.68; verbatimLongitude:135.83; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; eventDate:1960-10-29; individualCount:1; sex:male; preparations:pinned specimen, glued to a card; recordedBy:M. Sasakawa; identifiedBy:J. Salmela; institutionCode:OSAKA
Description
Male. Head black, vertex covered by pale setae, frons glabrous and face with scattered apical setae; face basally, close to scape, yellowish. Ocelli in a shallow triangle, median ocellus smallest. Clypeus with microtrichosity (pruinosity), elongated (about 1.7 times longer than basally wide). Scape yellowish and brownish, pedicel yellow and first flagellomere basally yellowish. Length ratio of pedicel:first flagellomere 0.24. Flagellomeres dark, palpus yellow.
Thorax dark-brown with pale setosity. Scutum shining, pleural sclerites with weak microtrichosity. Antepronotum yellow. Halter yellow. Femora yellow, bearing pale setae. Trochanters infuscated. Femora yellow, but mid and hind femora ventrobasally infuscated. Legs gradually darkening toward tarsi. Tibial spurs brownish. Length ratio of femur to tibia for fore, mid and hind legs: 0.77, 0.66, 0.66. Length ratio of tibia to basitarsus for fore, mid and hind legs: 1.06, 1.68, 1.68.
Apex of wing slightly infuscated. Bases of M1 and M2, M1+2, r-m, bM1+2, Rs, A1 and Sc bare, other veins setose. C exceeding tip of R5 36 % of the distance between R5 and M1. Sc ending in C at the level of Rs. Length ratio of M1+2:r-m = 1.19. Cu forking slightly beyond M end of r-m. Wing length 5.0 mm.
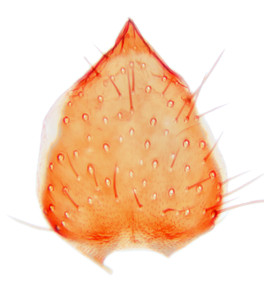
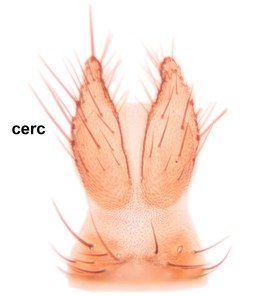
Abdomen dark-brown, tergites 2–4 laterodistally yellowish. 9th tergite elongated; cerci bearing two rows of combs, that are about equally wide, having ca. 45 stout setae (Fig.
Diagnosis
A large species with a vaguely infuscated wing apex, abdominal tergites 2–4 laterally yellowish and relatively long first flagellar segment (about 4-times the length of the pedicel). The ventral lobe of gonostylus bare, sinuous; in the closely related B. curta it is curved and bearing two stout setae. The apices of parameres with a conspicuous pair of horn-like outgrowths.
Etymology
The new species is named after Dr. Mitsuhiro Sasakawa, Japanese entomologist and the collector of the holotype. The name is a genitive.
Distribution
Known only from the type locality (Yoshino in Japan). The holotype male was collected at the end of October.
Taxon discussion
See above Boletina curta.
Boletina norokorpii , sp. n.
-
country: Finland; stateProvince:Ostrobothnia borealis pars borealis; verbatimLocality:Ylitornio, Tuorerommas Mire Conservation Area; verbatimLatitude:66.479; verbatimLongitude:24.757; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2012-7-2/8-6; habitat:old-growth boreal forest with a spring brook; individualCount:1; sex:male; catalogNumber:DIPT-JS-2016-0044; recordedBy:J. Salmela; identifiedBy:J. Salmela; institutionCode:ZMUT
Description
Male. Head black, vertex covered by pale setae, frons glabrous and face with scattered setae. Ocelli in a shallow triangle, median ocellus smallest. Clypeus not much longer than wide (about 1.2 times longer than basally wide). Scape and pedicel brownish, first and second flagellomeres yellowish, base of third flagellomere yellowish. Length ratio of pedicel:first flagellomere 0.36. Flagellomeres dark, palpus yellow.
Thorax dark-brown with pale setosity. Antepronotum yellow. Halter yellow. Femora yellow, bearing pale setae. Trochanters infuscated. Femora yellow. Legs gradually darkening toward tarsi. Tibial spurs brownish. Length ratio of femur to tibia for fore and hind legs: 0.93, 0.76. Length ratio of tibia to basitarsus of hind leg: 1.68.
Apex of wing slightly infuscated. Bases of M1 and M2, M1+2, r-m, bM1+2, Rs, A1 and Sc bare, other veins setose. C exceeding tip of R5 16 % of the distance between R5 and M1. Sc ending in C at the level of Rs. Sc2 present. Length ratio of M1+2:r-m = 1.14. Cu forking slightly beyond M end of r-m. Wing length 4.1 mm.
Abdomen dark-brown, tergites 2-4 laterodistally yellowish. 9th tergite elongated; cerci bearing two rows of combs, that are about equally wide, having 18 stout setae (Fig.
Diagnosis
A species very close to B. curta and B. sasakawai sp.n. The ventral lobe of the gonostylus of B. norokorpii sp.n. is curved, having no stout setae (setae present in B. curta; ventral lobe of the gonostylus in B. sasakawai sp.n. is sinuous). The caudal and proximal combs of the cerci are equally wide, having relatively a small number (18) of stout setae (over 40 in both B. curta and B. sasakawai sp.n.)
Etymology
The new species is named after Dr. Yrjö Norokorpi, Finnish forest researcher and former area manager at Parks & Wildlife Finland. The name is a genitive.
Distribution
So far known from SW Finnish Lapland only (Fig.
Ecology
The Finnish trapping site was an old-growth boreal forest characterised by vascular plants typical for base-rich soils, such as Paris quadrifolia and Calypso bulbosa.
Taxon discussion
The new species is very close to the eastern Palaearctic species B. curta and B. sasakawai sp.n. It is likely, however, that the eastern species are more related to each other than to B. norokorpii sp.n. For example, presence of Sc2 (absent in other species), shorter basal flagellar segments (1st flagellomere about 2.4 times longer than pedicel; in other species 3.1-4.2) and the small number (18; over 40 in other species) of stout setae of combs in the cerci separate the new species from the eastern Palaearctic taxa. We assume that both eastern Palaearctic species have restricted ranges in Japan, Far East Russia and neighbouring areas, whereas B. norokorpii sp.n. might have a widespread boreal range.
DNA barcoding
Holotype: BOLD Sample ID: DIPT-JS-2016-0044. BOLD Process ID: SCFI744-16. GenBank accession number: KY062991.
AATATTATATTTTATTTTTGGAGCTTGATCAGGAATAATTGGTACATCATTAAGAATTCTTATTCGTGCTGAATTAGGACACCCTGGAGCATTAATTGGAGATGATCAAATTTATAATGTTATTGTAACAGCTCATGCATTTGTAATAATTTTTTTTATAGTAATACCTATTATAATTGGAGGATTTGGTAATTGATTAATCCCTTTAATATTAGGAGCTCCTGATATAGCATTCCCTCGAATAAATAATATAAGATTTTGACTACTTCCTCCTTCATTAATATTACTTTTATCCAGAAGTTTAGTTGAAACAGGGGCTGGTACAGGTTGAACAGTGTACCCACCATTATCCTCAACAATTGCTCATGCAGGAGCATCTGTTGATTTAGCAATTTTTTCATTACATTTAGCAGGAATTTCTTCTATTTTAGGAGCTGTAAATTTTATTACTACAATTATTAATATACGAGCTCCTGGAATTACTTTTGAACGAATACCTCTTTTTGTATGATCAGTTTTAATTACAGCTATTTTATTATTATTATCTCTCCCAGTTTTAGCTGGAGCTATTACTATACTTTTAACAGACCGTAATTTAAATACATCATTTTTTGATCCTGCTGGAGGAGGAGATCCTATTTTATATCAACACTTATTC
The new species is assigned to the BIN BOLD:ADD1952, shared by no other specimens. In BOLD database the closest matches to this specimen are three Boletina lundbecki Lundström and four Boletina unassigned to taxonomic species (93,43 - 93,02 similarity).
Phronia sompio , sp. n.
-
country: Finland; stateProvince:Regio kuusamoensis; verbatimLocality:Salla, Värriö Strict Nature Reserve, Kuntasjoki; verbatimLatitude:67.749; verbatimLongitude:29.616; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2013-6-4/29; habitat:old-growth boreal riparian forest with seepages; individualCount:1; sex:male; catalogNumber:DIPT-JS-2014-0011; recordedBy:J. Salmela; identifiedBy:J. Salmela; institutionCode:ZMUT
-
country: Finland; stateProvince:Regio kuusamoensis; verbatimLocality:Salla, Värriö Strict Nature Reserve, Kuntasjoki; verbatimLatitude:67.750; verbatimLongitude:29.620; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2013-6-29/7-29; habitat:riparian forest with lush vegetation and large amount of decaying trees,; individualCount:1; sex:male; catalogNumber:DIPT-JS-2015-0101; recordedBy:J. Salmela; identifiedBy:J. Salmela; institutionCode:JES
-
country: Finland; stateProvince:Lapponia kemensis pars orientalis; verbatimLocality:Savukoski, Urho Kekkonen National Park, Tyyroja; verbatimLatitude:68.143; verbatimLongitude:28.574; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2014-7-1/8-5; habitat:riparian meadow, spring brook with abundant Palustriella mosses; individualCount:1; sex:male; catalogNumber:DIPT-JS- 2014-0404; recordedBy:J. Salmela; identifiedBy:J. Salmela; institutionCode:JES
Description
Male. Head brown, vertex covered by pale setae, frons glabrous. Ocelli in a line, central ocellus smallest, lateral ocelli close to eyes, their distance from eye less than their own width. Eyes pubescent. Palpi brown, bearing light setae. Length ratio of palpal segments 3-5: 3:4=0.88, 4:5=0.61. Penultimate segment 2.62 times as long as wide, last segment 4.67 times as long as wide. Third palpomere with a sensory pit in its base. Antennae brown, 16-segmented (scape, pedicel and 14 flagellomeres), pedicel and base of first flagellomere yellowish brown. Scape:pedicel length ratio 1.47. Flagellomeres cylindrical, length:width ratio of 1st flagellomere 2.27, 4th flagellomere 1.67 and apical flagellomere 1.90. Flagellomeres covered by dense light setosity, setae slightly curved, their length shorter than width of respective flagellomere.
Thorax generally brown, except scutum that has yellowish brown anterior corners. Scutum with mainly pale setosity, two stout and long posterodorsal setae are present just above scutellum. Mediotergite bare, other sclerites bearing setae. Scutellum with four stout marginal setae. Halteres pale, bearing weak light setae and setulae.
Wings hyaline, veins light brown. Bases of M1 and M2, M1+2, base of r-m, bM1+2, bRs and Sc bare, other veins setose. C slightly exceeding tip of R5. Sc ending free. Length ratio of M1+2:r-m = 1.03. Wing length 1.8 mm.
Coxae yellow, bearing dark setae, legs yellowish. Length ratio of femur to tibia for fore, mid and hind legs: 1.02, 1.0, 0.84. Length ratio of tibia to basitarsus for fore, mid and hind legs: 1.28, 1.6, 1.63. Anteroapical depressed area of the fore tibia ovate, having ca. 20 light setae arranged in a curved row. Ratio of apical width of tibia:length of longest tibial spur for fore, mid and hind legs: 0.37, 0.30, 0.29.
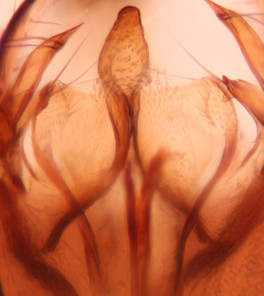
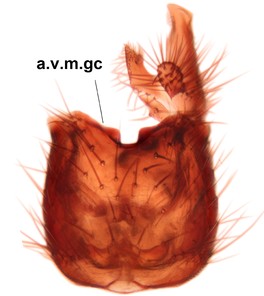
Abdominal tergites and sternites brown, bearing light setae. 9th tergite and cerci without peculiar characteristics. Ventroapical margin of gonocoxite with a marked median emargination (Fig.
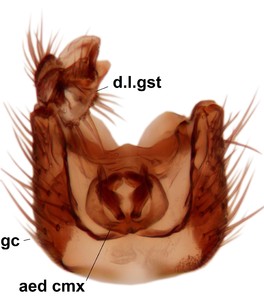
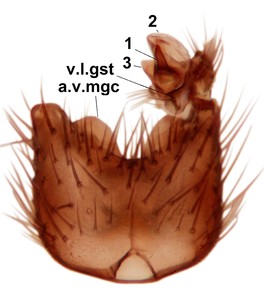


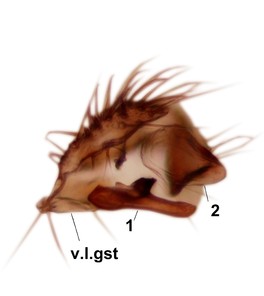
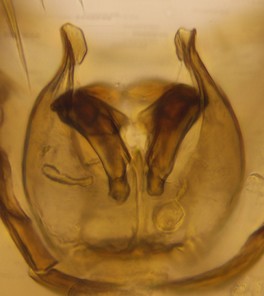
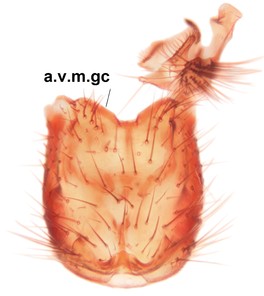
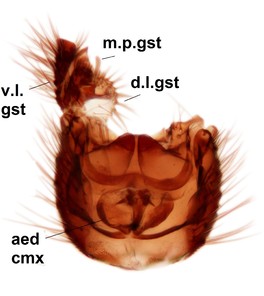
Phronia sompio Salmela sp.n., holotype male, hypopygium.
b: Hypopygium, ventral view. v.l.gst=ventral lobe of gonostylus, a.v.m.gc=ventroapical margin of gonocoxites, 1–3=projections of the mesial portion of gonostylus (see text).
c: Gonostylus, outer lateral view. m.p.gst=mesial portion of gonostylus.
d: Gonostylus, inner lateral view.
e: Gonostylus, dorsal view.
f: Aedeagal complex, dorsal view.
Diagnosis
A small species that is different from the known member of the genus. The male hypopygium has the following diagnostic characters: the ventroapical margin of the gonocoxites has a deep notch; the mesial projection of the gonostylus lacks comb-like structures but bears a prominent finger-like projection and a rounded, hyaline protrusion; the aedeagal complex is about as long as broad and is apically notched.
Etymology
The name of the new species refers to the old Forest Sami name of the region, Sompio, meaning large area bordered by aapamires. The biogeographical province of Lapponia kemensis pars orientalis, abbreviated as Lkor, is in Finnish "Sompion Lappi". The name is a noun in apposition.
Distribution
The species has been collected so far from three localities, all of these from eastern Finnish Lapland close to the Russian border. In fact, all of the collecting sites belong to the River Tuuloma catchment area east of the Maanselkä divide, so the waters finally flow to the Barents Sea in Russia.
Ecology
Collecting sites are small waterbodies (spring-fed headwater streams, spring brooks) surrounded by old-growth boreal forests.
Taxon discussion
The new species stands apart from all other Holarctic members of the genus.
DNA barcoding
BOLD Sample ID: DIPT-JS-2014-0011. BOLD Process ID: SCFI001-15.
BOLD Sample ID: DIPT-JS-2015-0101. BOLD Process ID: SCFI164-15.
BOLD Sample ID: DIPT-JS-2014-0404. BOLD Process ID: SCFI102-15.
Barcoding of the holotype and paratypes failed.
Phronia reducta , sp. n.
Nomenclature
Phronia annulata Winnertz,
-
country: Finland; stateProvince:Regio kuusamoensis; verbatimLocality:Salla, Iso Pyhätunturi; verbatimLatitude:66.776; verbatimLongitude:28.810; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2013-7-19/8-8; habitat:poor - intermediate rich sloping fen; individualCount:1; sex:male; catalogNumber:DIPT-JS-2015-0272; recordedBy:J. Salmela; identifiedBy:J. Salmela; institutionCode:ZMUT
-
country: Russia; stateProvince:Krasnoyarsk region; verbatimLocality:Tungussko-Chunsky District, village Vanavary; verbatimLatitude:60.33; verbatimLongitude:102.30; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:sweep net; eventDate:1972-7-26; habitat:swampy forest; individualCount:1; sex:male; preparations:slide mounted; catalogNumber:1386 (3); recordedBy:G.P. Ostroverkhova; identifiedBy:J. Salmela; institutionCode:TSU
Description
Male. Head dark-brown, vertex covered by pale setae, frons glabrous. Ocelli in a line, central ocellus slightly smaller than laterals; lateral ocelli close to eyes, their distance from eye less than their own width. Eyes pubescent. Palpi brown, bearing light setae. Length ratio of palpal segments 3–5: 3:4=0.83, 4:5=0.69. Penultimate segment 3.6 times as long as wide, last segment 5.3 times as long as wide. Third palpomere with a sensory pit in its base. Antennae brown, 16-segmented (scape, pedicel and 14 flagellomeres), base of pedicel and base of first flagellomere yellowish brown. Scape:pedicel length ratio 1.60. Flagellomeres cylindrical, length:width ratio of 1st flagellomere 2.86, 4th flagellomere 1.75 and apical flagellomere 3.0. Flagellomeres covered by dense light setosity, setae slightly curved, their length shorter than width of respective flagellomere.
Thorax generally dark-brown, except scutum that has yellowish anterior corners. Scutum with mainly pale setosity. Mediotergite bare, other sclerites bearing setae. Scutellum with four stout setae. Halteres pale, bearing weak light setae and setulae.
Wings hyaline, veins brown. Bases of M1 and M2, M1+2, base of r-m, bM1+2, base of Rs and Sc bare, other veins setose. C exceeds tip of R5 very slightly. Sc ending free. Length ratio of M1+2:r-m = 1.18. Wing length 3.1 mm.
Coxae yellow, bearing pale setae, legs yellowish, except femora ventrobasally and apices of hind femora infuscated. Length ratio of femur to tibia for fore, mid and hind legs: 0.95, 0.99, 0.82. Length ratio of tibia to basitarsus for fore, mid and hind legs: 1.03, 1.34, 1.60. Anteroapical depressed area of the fore tibia ovate, having ca. 19 light setae arranged in a slightly curved row. Ratio of apical width of tibia:length of longest tibial spur for fore, mid and hind legs: 0.39, 0.27, 0.24.
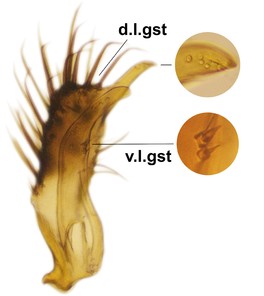
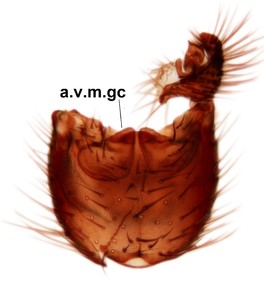
Abdominal tergites and sternites brown, bearing light setae. 9th tergite and cerci normal for the genus (Fig.

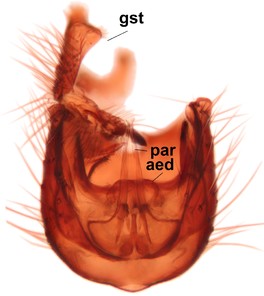


Phronia reducta Salmela sp.n., holotype male, hypopygium.
b: Hypopygium, dorsal view. gst=gonostylus, par=parameres, aed=aedeagus.
c: Hypopygium, ventral view. a.v.m.gc=ventroapical margin of gonocoxites.
d: Gonostylus, outer lateral view. d.l.gst=dorsal lobe of gonostylus. 1-4, outgrowths of the gonostylus, see text.
e: Gonostylus, inner lateral view. v.l.gst=ventral lobe of gonostylus.
f: Aedeagus and parameres, dorsal view. Insert shows apices of parameres in lateral view.
Diagnosis
The new species is close of P. braueri Dziedzicki but differs in the following features; the apices of the parameres are non-setose (the setae here are long in P. braueri), the internal outgrowth of the ventral lobe of the gonostylus is curved and apically notched (not curved or notched in P. braueri), and the ventral lobe of the gonostylus also has a narrow club-like basal projection (wedge-shaped and widest basally in P. braueri).
Etymology
The name of the new species (Latin reducta, reduced, an adjective) is referring to the non-setose apices of the male parameres.
Distribution
Apparently a boreal species, hitherto known from NE Finnish Lapland and Siberia, Central Russia (Fig.
Ecology
The species occurs in sloping fens and swampy forests. The Finnish collecting site (sloping fen) was close to a pine and spruce dominated pristine boreal forest.
Taxon discussion
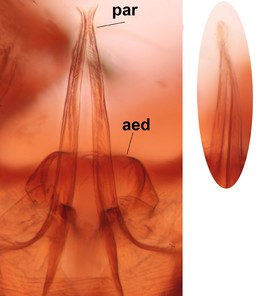
The new species was illustrated for the first time by

Phronia braueri Dziedzicki, male hypopygium. The specimen (MYCE-JS-2013-0014, JES) is collected from Finland, Lkor: Savukoski, Joutenoja, 16.8.-18.9.2012, J. Salmela leg. Please see Fig.
b: Hypopygium, ventral view.
c: Gonostylus, inner lateral view.
d: Gonostylus, outer lateral view.
There are two questionable older names of P. braueri, namely P. annulata Winnertz and P. vittata Winnertz (
DNA barcoding
Holotype male: BOLD Sample ID: DIPT-JS-2015-0272. BOLD Process ID: SCFI741-16. GenBank accession number: KY062992.
AATTTTATATTTTATTTTTGGAGCTTGATCTGGAATAGTGGGAACTTCTCTTAGAATTATTATTCGGACTGAATTAGGACATCCAGGAGCATTAATTGGTAATGACCAAATTTATAATGTTATTGTTACAGCTCATGCTTTTATTATAATTTTTTTTATAGTTATACCTATTATAATTGGAGGATTTGGAAATTGATTAGTTCCACTAATACTAGGAGCCCCTGATATAGCTTTTCCTCGAATAAATAATATAAGATTTTGGTTATTACCTCCTTCTCTTACATTATTACTTTCTAGAAGTTTAGTAGAAGCAGGGGCTGGAACTGGTTGAACAGTTTACCCTCCCCTTTCTTCAACTATTGCTCATGCTGGCGCATCAGTTGATTTAGCTATTTTTTCTTTACATTTAGCAGGTATTTCATCAATTTTAGGGGCAGTTAATTTTATTACTACCATTATTAATATACGAGCTCCTGGAATCACTTTTGATCGTTTACCTTTATTTGTTTGATCTGTTCTTATTACAGCAGTATTACTATTATTATCTTTACCCGTATTAGCAGGAGCTATTACTATACTATTAACAGACCGAAATCTTAATACTTCATTTTTTGACCCTGCAGGGGGAGGAGATCCTATTTTATACCAACATTTATTT
The holotype male is the only member of the BIN BOLD:ADD3565. This specimen has no very close matches in BOLD database. The closest matches are 44 Phronia specimens, whose similarities to the new species range between 96,74 - 96,01. One of these specimens is assigned to P. braueri, the sister species of P. reducta sp.n. That P. braueri specimen is collected from Norway and was identified by J. Kjaerandsen (unpublished record).
Phronia prolongata , sp. n.
-
country: Finland; stateProvince:Lapponia inarensis; verbatimLocality:Inari, Muotkatunturi Wilderness Area, Kielajoki; verbatimLatitude:69.146; verbatimLongitude:26.292; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2014-6-26/8-5; habitat:lush and swampy riparian birch forest; individualCount:1; sex:male; catalogNumber:DIPT-JS-2015-0215; recordedBy:E. Rundgren; identifiedBy:J. Salmela; institutionCode:ZMUT
-
country: Norway; stateProvince:Finnmark; verbatimLocality:Alta, Goppaelva; verbatimLatitude:70.027; verbatimLongitude:23.394; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:sweep net; eventDate:2010-6-13; individualCount:1; sex:male; catalogNumber:MYCFI183-11; recordedBy:Finnmarksprosjektet; identifiedBy:J. Salmela, G. Söli; institutionCode:NHMO
-
country: Norway; stateProvince:Finnmark; verbatimLocality:Alta, Goppaelva; verbatimLatitude:70.027; verbatimLongitude:23.394; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:sweep net; eventDate:2010-6-13; individualCount:1; sex:male; catalogNumber:MYCFI184-11; recordedBy:Finnmarksprosjektet; identifiedBy:J. Salmela, G. Söli; institutionCode:NHMO
-
country: Germany; stateProvince:Bavaria; verbatimLocality:Allgäu, Oberstdorf, Schochen; verbatimElevation:2032 m; verbatimLatitude:47.3936; verbatimLongitude:10.3692; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2014-6-6/21; habitat:Blaugras-Horstseggenrasen; individualCount:1; sex:male; catalogNumber:BC-ZSM-DIP-22552-E10; recordedBy:D. Doczkal, S. Schmidt & J. Voith; identifiedBy:J. Salmela; institutionCode:ZSM
-
country: Canada; stateProvince:British Columbia; verbatimLocality:Vancouver Island; verbatimLatitude:49.044; verbatimLongitude:-125.684; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:sweep net; eventDate:2014-6-30; habitat:old growth temperate rain forest; individualCount:1; sex:female; catalogNumber:BIOUG21868-H06; recordedBy:BIObus; identifiedBy:BOLD ID engine; institutionCode:BIOUG
Description
Male. Head brown, vertex covered by pale setae, frons glabrous. Ocelli arranged in a very shallow triangle, central ocellus slightly smaller than laterals; lateral ocelli close to eyes, their distance from eye less than their own width. Eyes pubescent. Palpi brown, bearing light setae. Length ratio of palpal segments 3–5: 3:4=0.92, 4:5=0.62. Penultimate segment 3.51 times as long as wide, last segment 10.0 times as long as wide. Third palpomere with a sensory pit in its base. Antennae brown, 16-segmented (scape, pedicel and 14 flagellomeres). Scape:pedicel length ratio 1.28. Flagellomeres cylindrical, length:width ratio of 1st flagellomere 3.0, 4th flagellomere 2.58 and apical flagellomere 3.2. Flagellomeres covered by dense light setosity, setae slightly curved, their length shorter than width of respective flagellomere.
Thorax generally brown, scutum dorsally dark-brown. Scutum with mainly pale setosity, including the two stout and long dorso-posterior setae above scutellum. Mediotergite bare, other sclerites bearing setae. Scutellum with four stout marginal setae. Halteres pale, bearing weak light setae and setulae.
Wings hyaline, veins light brown. Bases of M1 and M2, M1+2, r-m, bM1+2, bRs and apex of Sc bare, other veins setose. C slightly exceeding tip of R5. Sc ending free. Length ratio of M1+2:r-m = 1.29. Wing length 3.2-3.5 mm.
Coxae and legs yellowish brown, bearing dark setae. Length ratio of femur to tibia for fore and mid legs (hind legs are missing from the holotype, ratios of that leg are form the German paratype): 0.93, 0.9, 0.76. Length ratio of tibia to basitarsus for fore and mid legs: 0.96, 1.21, 1.5. Anteroapical depressed area of the fore tibia ovate, having numerous light setae over the area. Ratio of apical width of tibia:length of longest tibial spur for fore and mid legs: 0.67, 0.33, 0.23.
Abdomen. 9th tergite and cerci as in Fig.

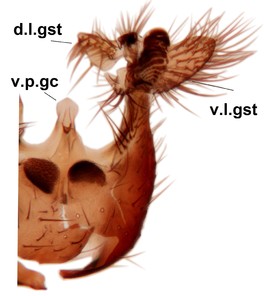
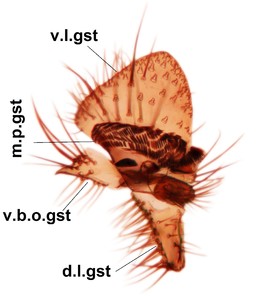
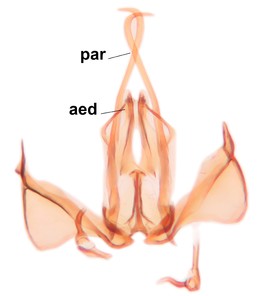

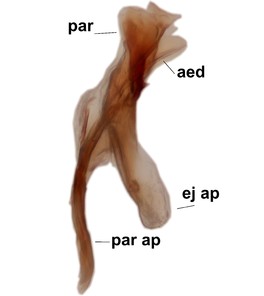
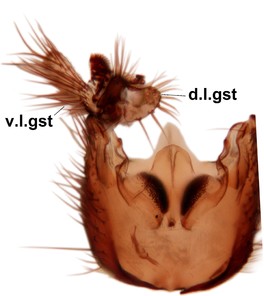
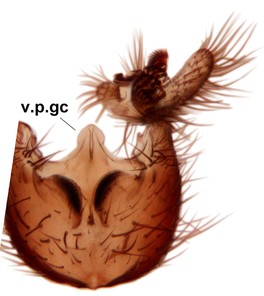
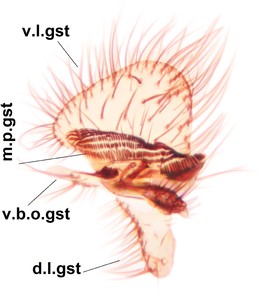
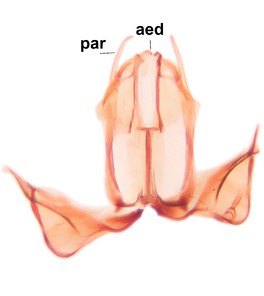
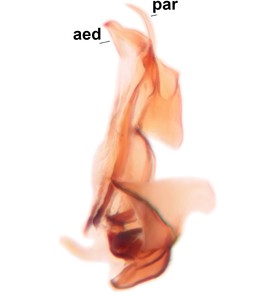
Phronia prolongata Salmela sp.n., holotype male, hypopygium.
b: Hypopygium, dorsal view.
c: Hypopygium, ventral view. d.l.gst=dorsal lobe of gonostylus, v.l.gst=ventral lobe of gonostylus, v.p.gc=ventrocaudal projection of gonocoxites.
d: Gonostylus, inner lateral view. m.p.gst=mesial portion of gonostylus, v.b.o.gst=ventrobasal outgrowth of v.l.gst.
e: Aedeagal complex, dorsal view. par=parameres, aed=aedeagus.
f: Aedeagal complex, lateral view.
Phronia exigua Zetterstedt, male hypopygium. The specimen (MYCE-JS-2013-0010, JES) is collected from Finland, Lkor: Savukoski, Joutenoja, 16.8.-18.9.2012, J. Salmela leg. Please see Fig.
b: Hypopygium, dorsal view.
c: Hypopygium, ventral view.
d: Gonostylus, inner lateral view.
e: Aedeagal complex, dorsal view.
f: Aedeagal complex, lateral view.
Female. In general, similar to male. Antennae dark except scape, pedicel and base of 1st flagellomere yellowish brown. Scape:pedicel length ratio 1.54. Length:width ratio of 1st flagellomere 3.1, 4th flagellomere 2.14 (apical flagellomeres broken off). Length ratio of M1+2:r-m = 1.58. Wing length 3.5 mm.
Diagnosis
Phronia prolongata sp.n. is a closely related species of P. exigua (Zetterstedt, see Fig.
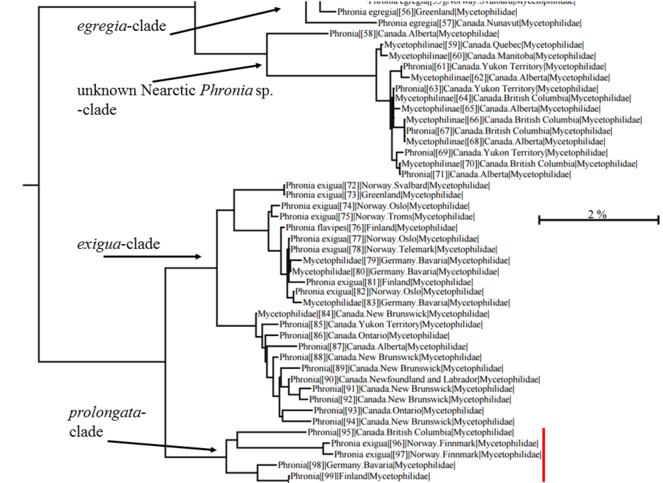
NJ-tree of Phronia prolongata sp.n. and related taxa based on COI (mtDNA) sequences and K2P distances (data from BOLD database). 95–98 represent P. prolongata sp.n. specimens studied here; the species displays a notable intraspecific variation of the barcoding gene COI, but all specimens are considered conspecific. The Canadian specimen is a female, and its identification is solely based on the COI sequence; Nearctic male specimens should be seen in order to validate the taxonomic assignment proposed here.
Etymology
The name (Latin prolongata, an adjective) of the new species refers to the elongated, prolonged parameres of the male hypopygium.
Distribution
The new species has a Holarctic range, it is known from Canada (British Columbia), Norway, Finland and Germany (Fig.
Ecology
The Finnish collecting site was a swampy riparian birch forest and in Germany the species was collected from an alpine meadow.
Taxon discussion
The new species belongs to a group of species clastered with P. exigua, all sharing a beaked, setose ventral lobe of the gonostylus (ventrobasal outgrowth of the ventral lobe of gonostylus) and a row of ventral setae on the hind tibia (
DNA barcoding
BOLD Sample ID: DIPT-JS-2015-0215. BOLD Process ID: SCFI251-15. GenBank accession number: KY062993.
AATTTTATACTTTATTTTTGGTGCTTGATCTGGAATAGTAGGAACTTCCCTAAGAATTATTATTCGTGCTGAACTTGGTCATCCAGGAGCATTGATTGGAAATGATCAAATTTATAATGTAATTGTTACTGCTCATGCTTTCATTATAATTTTTTTTATAGTTATGCCCATTATAATTGGTGGGTTTGGTAACTGACTTGTCCCATTGATATTGGGGGCCCCTGATATAGCTTTTCCTCGAATAAATAATATAAGTTTCTGATTATTGCCTCCCTCATTAACACTTCTTCTTTCAAGAAGTTTAGTCGAAGCTGGGGCTGGTACAGGTTGAACTGTTTATCCCCCTCTTTCTTCTACTATTGCTCACGCAGGATCTTCTGTTGATCTAGCTATTTTTTCTCTTCATTTAGCTGGTATTTCTTCAATTTTAGGGGCGGTAAATTTTATCACAACTATTATTAATATACGAGCTCCAGGAATTTCCTTTGATCGTTTACCTTTATTTGTTTGATCTGTTCTTATTACTGCTGTATTGCTTCTTTTATCGCTACCAGTTTTAGCTGGGGCTATTACTATACTTTTAACTGATCGAAATTTAAACACATCTTTCTTTGACCCTGCCGGAGGGGGGGACCCTATTCTTTATCAACATTTATTT
The similarity of COI sequences between the new species and P. exigua range between 95.57 and 94.8, and between the new species and P. egregia 89.98-87.86. The new species displays a notable intraspecific variation: the Canadian non-type female has 97.06 similarity compared to the holotype and the two Norwegian specimens have 96.6 similarity. Interestingly the similarity of the holotype and German paratype is 98.94, and these two are classified to the same Barcode index number (BIN) by the BOLD (BOLD:ACW2188), shared by no other specimens. The new species is, however, monophyletic (Fig.
Phronia elegantula
Nomenclature
-
country: Finland; stateProvince:Ostrobothnia kajanensis; verbatimLocality:Sotkamo, Aarreniemi; eventDate:1964-8-11; individualCount:1; sex:male; recordedBy:A.V.V. Mikkola; identifiedBy:W. Hackman; institutionCode:MZHF
-
country: Finland; stateProvince:Regio kuusamoensis; verbatimLocality:Kuusamo, Juuma, Jäkälävuoma; eventDate:1964-8-21; individualCount:1; sex:male; recordedBy:R. Tuomikoski, K. Mikkola; identifiedBy:W. Hackman; institutionCode:MZHF
-
country: Finland; stateProvince:Lapponia kemensis pars orientalis; verbatimLocality:Pelkosenniemi, Luiron suot Mire Conservation Area, Sudenvaaranaapa; verbatimLatitude:67.1900; verbatimLongitude:27.6352; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2015-7-31/9-29; habitat:rich birch fen; individualCount:1; sex:male; catalogNumber:DIPT-JS-2016-0166; recordedBy:J. Salmela; identifiedBy:J. Salmela; institutionCode:JES
-
country: Finland; stateProvince:Lapponia kemensis pars orientalis; verbatimLocality:Pelkosenniemi, Luiron suot Mire Conservation Area, Sudenvaaranaapa; verbatimLatitude:67.1900; verbatimLongitude:27.6352; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2015-7-31/9-29; habitat:rich birch fen; individualCount:1; sex:male; catalogNumber:DIPT-JS-2016-0167; recordedBy:J. Salmela; identifiedBy:J. Salmela; institutionCode:JES
-
country: Norway; stateProvince:Buskerud; verbatimLocality:Kongsberg, Skollenborg, Labro; verbatimLatitude:59.6184; verbatimLongitude:9.6774; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:sweep net; eventDate:2008-9-28; individualCount:1; sex:male; catalogNumber:NHMO_MYC00025; recordedBy:L.O. Hansen; identifiedBy:J. Salmela; institutionCode:NHMO
-
country: Norway; stateProvince:Buskerud; verbatimLocality:Kongsberg, Skollenborg, Labro; verbatimLatitude:59.6184; verbatimLongitude:9.6774; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:sweep net; eventDate:2008-9-28; individualCount:1; sex:male; catalogNumber:NHMO_MYC00026; recordedBy:L.O. Hansen; identifiedBy:J. Salmela; institutionCode:NHMO
-
country: Germany; stateProvince:Bavaria; verbatimLocality:Nationalpark Bayerischer Wald, 11.3 km N of Grafenau; verbatimElevation:842 m; verbatimLatitude:48.950; verbatimLongitude:13.421; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2012-8-25/9-3; habitat:conifer-dominated mountain forest; individualCount:1; sex:male; catalogNumber:BIOUG08254-E11; recordedBy:G. Sellmayer; identifiedBy:J. Salmela; institutionCode:ZSM
-
country: Germany; stateProvince:Bavaria; verbatimLocality:Nationalpark Bayerischer Wald, 11.3 km N of Grafenau; verbatimElevation:842 m; verbatimLatitude:48.950; verbatimLongitude:13.421; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2012-8-25/9-3; habitat:conifer-dominated mountain forest; individualCount:1; sex:male; catalogNumber:BIOUG08259-G06; recordedBy:G. Sellmayer; identifiedBy:J. Salmela; institutionCode:ZSM
-
country: Germany; stateProvince:Bavaria; verbatimLocality:Nationalpark Bayerischer Wald, 11.3 km N of Grafenau; verbatimElevation:842 m; verbatimLatitude:48.950; verbatimLongitude:13.421; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2012-8-25/9-3; habitat:conifer-dominated mountain forest; individualCount:1; sex:female; catalogNumber:BIOUG08318-G10; recordedBy:G. Sellmayer; identifiedBy:J. Salmela; institutionCode:ZSM
-
country: Germany; stateProvince:Bavaria; verbatimLocality:Nationalpark Bayerischer Wald, 11.3 km N of Grafenau; verbatimElevation:842 m; verbatimLatitude:48.950; verbatimLongitude:13.421; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2012-8-25/9-3; habitat:conifer-dominated mountain forest; individualCount:1; sex:female; catalogNumber:BIOUG08251-F07; recordedBy:G. Sellmayer; identifiedBy:J. Salmela; institutionCode:ZSM
-
country: Germany; stateProvince:Bavaria; verbatimLocality:Nationalpark Bayerischer Wald, 11.3 km N of Grafenau; verbatimElevation:842 m; verbatimLatitude:48.950; verbatimLongitude:13.421; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2012-8-25/9-3; habitat:conifer-dominated mountain forest; individualCount:1; sex:female; catalogNumber:BIOUG08217-B09; recordedBy:G. Sellmayer; identifiedBy:J. Salmela; institutionCode:ZSM
-
country: Germany; stateProvince:Bavaria; verbatimLocality:Nationalpark Bayerischer Wald, 11.3 km N of Grafenau; verbatimElevation:842 m; verbatimLatitude:48.950; verbatimLongitude:13.421; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2012-8-25/9-3; habitat:conifer-dominated mountain forest; individualCount:1; sex:female; catalogNumber:BIOUG08218-G07; recordedBy:G. Sellmayer; identifiedBy:J. Salmela; institutionCode:ZSM
-
country: Germany; stateProvince:Bavaria; verbatimLocality:Nationalpark Bayerischer Wald, 11.3 km N of Grafenau; verbatimElevation:842 m; verbatimLatitude:48.950; verbatimLongitude:13.421; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2012-8-25/9-3; habitat:conifer-dominated mountain forest; individualCount:1; sex:female; catalogNumber:BIOUG08217-C03; recordedBy:G. Sellmayer; identifiedBy:J. Salmela; institutionCode:ZSM
-
country: Germany; stateProvince:Bavaria; verbatimLocality:Nationalpark Bayerischer Wald, 11.3 km N of Grafenau; verbatimElevation:842 m; verbatimLatitude:48.950; verbatimLongitude:13.421; verbatimCoordinateSystem:decimal degrees; verbatimSRS:WGS84; samplingProtocol:Malaise trap; eventDate:2012-8-2/12; habitat:conifer-dominated mountain forest; individualCount:1; sex:female; catalogNumber:BIOUG08211-A12; recordedBy:G. Sellmayer; identifiedBy:J. Salmela; institutionCode:ZSM
Description
Male. Head dark-brown, vertex covered by dark setae, frons glabrous and face anteriorly with small setae. Ocelli arranged in a line, central ocellus smaller than laterals; lateral ocelli close to eyes, their distance from eye less than their own width. Eyes pubescent. Palpi yellowish-brown, bearing light setae. Length ratio of palpal segments 3–5: 3:4=0.98, 4:5=0.59. Penultimate segment 2.94 times as long as wide, last segment 8.2 times as long as wide. Third palpomere with a sensory pit in its base. Antennae brown, 16-segmented (scape, pedicel and 14 flagellomeres); scape, pedicel and basal half of first flagellomere yellowish. Scape with a prominent dorsal seta, about as long as first flagellomere. Scape:pedicel length ratio 1.33. Flagellomeres cylindrical, length:width ratio of 1st flagellomere 2.98, 4th flagellomere 1.75 and apical flagellomere 2.95. Flagellomeres covered by dense light setosity, setae slightly curved, their length shorter than width of respective flagellomere.
Thorax generally brown. Scutum dorsally with three dark stripes, that are almost confluent; the stripes are separated by very narrow yellowish gaps; anterolateral corners yellowish. Scutum with mainly pale setosity. Mediotergite bare, other sclerites bearing setae. Scutellum with four stout setae. Halteres pale, bearing weak light setae and setulae.
Wings hyaline, veins light brown. Bases of M1 and M2, M1+2, r-m, bM1+2, base of Rs and apex of Sc bare, other veins setose. C very slightly exceeding tip of R5. Sc ending free. Length ratio of M1+2:r-m = 1.29. Wing length 2.2 mm.
Coxae and legs yellow, apices of mid and hind femora sligthly infuscated, bearing dark setae. Length ratio of femur to tibia for fore, mid and hind legs: 0.99, 0.97, 0.79. Length ratio of tibia to basitarsus for fore, mid and hind legs: 1.08, 0.97, 0.8. Anteroapical depressed area of the fore tibia ovate, having ca. 20 light in a row. Ratio of apical width of tibia:length of longest tibial spur for fore, mid and hind legs: 0.35, 0.28, 0.25.
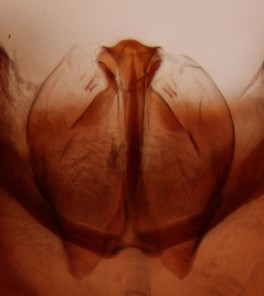
Abdomen mostly dark brown, but first, second and third tergites caudolaterally yellowish; these yellow areas are most extensive in second and third tergite. Sternum of second and third segments yellowish. Hypopygium dark brown. Ventroapical margin of gonocoxite with a wide and shallow median emargination, with a moderate median peak (Fig.
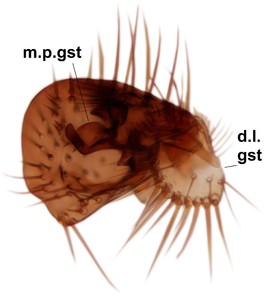
Phronia elegantula Hackman, male hypopygium, DIPT-JS-2016-0166.
b: Hypopygium, ventral view. a.v.m.gc=ventroapical margin of gonocoxites.
c: Gonostylus, inner lateral view.
d: Aedeagal complex, dorsal view.
Female. Similar to male. Antennae dark except scape, pedicel and base of 1st flagellomere yellowish brown. Scape:pedicel length ratio 1.32. Length:width ratio of 1st flagellomere 3.9, 4th flagellomere 2.80, apical flagellomere 2.5. Length ratio of M1+2:r-m = 1.84. Wing length 2.2 mm.
Diagnosis
A Phronia species with a yellowish pattern on the abdominal tergites 1–3. The ventral lobe of the gonostylus is rounded and at its widest basally. The mesial projections are finger-like and the inner lamella of the ventral lobe of the gonostylus bears a tuft of setae. The species is somewhat close to P. elegans Dziedzicki and P. signata Winnertz, that have similarly shaped ventral lobe of the gonostylus; P. elegantula can be distinguished from these due to differences in the structure of the aedeagus, the ventral lobe of gonostylus and the mesial portion of the gonostylus.
Distribution
A European species. The species was described from eastern Finland (Ok: Sotkamo and Ks: Kuusamo) and has been later recorded from southern and northern parts of the country (J. Jakovlev, unpublished). The species has been found from Russian Karelia (
Ecology
Sampling sites are coniferous forests, mixed forests and wetlands.
Taxon discussion
Phronia elegantula is somewhat similar to P. signata and P. elegans, and has the same yellowish anterolateral corners to the scutum as well as a rotund ventral lobe of the gonostylus. However the abdomen of P. elegans is dark brown as opposed to some yellowish colouration on abdominal tergites 1–3 of P. elegantula. Phronia signata have only moderately emarginated ventroapical margins of the gonocoxites, whereas this character is much more conspicuous in P. elegantula. Phronia signata has ca. 14 setae on the ventral edge of the ventral lobe of gonostylus (see e.g.
DNA barcoding
BOLD Sample ID: DIPT-JS-2016-0166. BOLD Process ID: SCFI751-16. GenBank accession number: KY200862. BOLD Sample ID: DIPT-JS-2016-0167. BOLD Process ID: SCFI752-16. GenBank accession number: KY200863. The sequence provided here is from DIPT-JS-2016-0166.
TATTTTATATTTCATTTTTGGTGCTTGATCTGGTATAGTAGGTACTTCTTTAAGAATCATTATTCGAACAGAATTAGGACACCCTGGAGCCTTAATTGGAAATGATCAAATTTATAATGTTATTGTTACTGCTCACGCTTTTATTATAATTTTTTTTATAGTTATACCAATTATAATTGGAGGATTCGGTAATTGATTAGTTCCACTAATATTAGGAGCTCCAGATATAGCTTTCCCTCGAATAAATAATATAAGTTTTTGACTTTTACCACCATCTTTAACCTTATTACTTTCTAGTAGCTTAGTAGAAGCAGGGGCTGGAACAGGATGAACTGTTTATCCCCCTTTATCATCTACAATTGCCCATGCAGGAGCCTCAGTTGATTTAGCTATCTTTTCTTTACATTTAGCAGGTATTTCTTCTATTTTAGGAGCAGTAAATTTTATTACAACAATTATTAATATACGGGCCCCAGGAATTACTTTTGACCGAATACCATTATTTGTTTGATCGGTATTAATTACAGCAGTTCTTCTATTACTTTCTCTACCAGTTTTAGCTGGAGCTATTACTATATTATTAACAGATCGAAATTTAAATACCTCATTTTTTGACCCTGCCGGAGGAGGAGATCCCATTTTATATCAACACTTATTT
All studied specimens belong to the BIN BOLD:ACJ2889, and their similarities range between 99.69 and 98.78 (average 99.46). The nearest specimens in BOLD database belong to P. disgrega Dziedzicki, being 90.98 % similar to P. elegantula. DNA barcode and associated data of the German paratypes and female specimens is available from the BOLD Public data portal.
Acknowledgements
We thank Matti Mäkilä, Eerikki Rundgren, Arto Saikkonen and Ari Kekki for their help in the field work. The second author work was financially supported by a grant of the Romanian Ministry of National Education, CNCS–UEFISCDI–nr. PN–II–ID–PCE–2012–4–0595. Barcode sequences were obtained at the Canadian Centre for DNA Barcoding based in the Biodiversity Institute of Ontario at the University of Guelph. Their work was supported by funding from the Government of Canada through Genome Canada and the Ontario Genomics Institute in support of the International Barcode of Life Project. Finnish Barcode of Life (FinBOL), especially Marko Mutanen and Mikko Pentinsaari (University of Oulu) are also thanked for their help in the barcoding process. Elena Subbotina (TSU) commented upon Phronia material examined by Ostroverkhova and she helped us to select the paratype of P. reducta sp.n. Dieter Doczkal (ZSM), Valerie Levesque-Beaudin (BIOUG), Rikio Matsumoto (OSAKA), Alexei Polevoi (FRIP), Pekka Vilkamaa (MZHF) and Geir Søli (NHMO) kindly loaned specimens for our disposal. Comments by Peter J. Chandler, Olavi Kurina and Vladimir Blagoderov improved the manuscript. English text was checked by Peter Boardman (Shrewsbury).
Author contributions
JS wrote the ms. L-PK took all photos on male hypopygia. JS is the author of all new species except Boletina sasakawai and B. norokorpii, that are coauthored: L-PK suggested that a Finnish specimen (holotype of B. norokorpii) may be a distinct species, and after examining type material of B. curta JS agreed this opinion.
References
- Pesky gnats: Ridding dipteran classification of the “Nematocera”.Studia dipterologica13(1):3‑9.
- Phylogeny of the Mycetophiliformia, with proposal of the subfamilies Heterotrichinae, Ohakuneinae, and Chiletrichinae for the Rangomaramidae (Diptera, Bibionomorpha).Zootaxa1535:1‑92.
- Systematics and Phylogeny of Leptomorphus Curtis (Diptera: Mycetophilidae).Zootaxa3549:1‑117.
- Areview of the British Phronia Winnertz and Trichonta Winnertz (Dipt., Mycetophilidae).Entomologist’s Monthly Magazine128:237‑254.
- deWaard J, Ivanova N, Hajibabaei M, Hebert PN (2008) Assembling DNA Barcodes. Methods in Molecular Biology. https://doi.org/10.1007/978-1-59745-548-0_15
- Revue des espèces européennes du genre Phronia Winnertz avec la description des deux genres nouveaux: Macrobrachius et Megophthalmidia.Horae Soc. Ent. Ross.23:404‑532.
- Literatura Taxonomica Dipterorum (1758–1930).Backhuys Publishers,Leiden,450pp.
- Catalog of the Keroplatidae of the World (Insecta: Diptera).Bishop Museum Bulletin in Entomology13:1‑178. URL: http://pbs.bishopmuseum.org/pdf/BE13-kero-catr.pdf
- A revision of the Nearctic species of the genus Phronia (Diptera: Mycetophiilidae).Transactions of the American Entomological Society101:227‑318.
- Testing the Global Malaise Trap Program – How well does the current barcode reference library identify flying insects in Germany?Biodiversity Data Journal4:e10671. https://doi.org/10.3897/bdj.4.e10671
- Nuove specie di Ditteri del Muzeo Zoologico di Torino.Bollettino dei Musei di Zoologia ed Anatomia Comparata della Regia Universita di Torino5(84):1‑4.
- New species of the genus Phronia Winnertz (Diptera, Mycetophilidae) from Eastern Fennoscandia and notes on the synonymies in this genus.Notulae entomologicae50:41‑60.
- Hackman W, Laštovka P, Matile L, Väisänen R (1988) Family Mycetophilidae (Fungivoridae). In: Soós A, Papp L (Eds) Catalogue of Palaearctic Diptera. Volume 3. Ceratopogonidae - Mycetophilidae.Akadémiai Kiadó,Budapest.
- Biological identifications through DNA barcodes.Proceedings of the Royal Society B: Biological Sciences270(1512):313‑321. https://doi.org/10.1098/rspb.2002.2218
- Counting animal species with DNA barcodes: Canadian insects.Philosophical Transactions of the Royal Society B: Biological Sciences371(1702):20150333. https://doi.org/10.1098/rstb.2015.0333
- Mycetophilidae (Bolitophilinae, Ditiomyiinae, Diadocidiinae, Keroplatinae, Sciophilinae and Manotinae). Diptera, Nematocera. Handbooks for Identification of British Insects.Royal Entomological Society of London,109pp.
- Do avian predators select for seasonal polyphenism in the European map butterfly Araschnia levana (Lepidoptera: Nymphalidae)?Biological Journal of the Linnean Society106(4):737‑748. https://doi.org/10.1111/j.1095-8312.2012.01922.x
- Fungus gnats (Diptera: Sciaroidea) associated with dead wood and wood growing fungi: new rearing data from Finland and Russian Karelia and general analysis of known larval microhabitats in Europe.Entomologica Fennica22:157‑189.
- Two new species of the genus Phronia Winnertz (Diptera: Mycetophilidae) from Finland and Russian Karelia.Entomologica Fennica19:199‑206.
- Recent noteworthy findings of fungus gnats from Finland and northwestern Russia (Diptera: Ditomyiidae, Keroplatidae, Bolitophilidae and Mycetophilidae).Biodiversity Data Journal2:e1068. https://doi.org/10.3897/bdj.2.e1068
- The Mycetophila ruficollis Meigen (Diptera, Mycetophilidae) group in Europe: elucidating species delimitation with COI and ITS2 sequence data.ZooKeys508:15‑51. https://doi.org/10.3897/zookeys.508.9814
- Checklist of Nordic fungus gnats (Diptera: Bolitophilidae, Diadocidiidae, Ditomyiidae, Keroplatidae, Mycetophilidae and Sciarosoma). Latest update 13.5.2016.http://sciaroidea.info/node/48341#. Accessed on: 2016-10-15.
- Annotated checklist of fungus gnats from Sweden (Diptera: Bolitophilidae, Diadocidiidae, Ditomyiidae, Keroplatidae and Mycetophilidae).Insect Systematics & Evolution, Supplement65:1‑165.
- Redescription of Sciophila nitens Winnertz (Diptera: Mycetophilidae) with a new synonymization.Entomologica Fennica15:193‑197.
- Kurina O (2008) Sciaroidea excl. Sciaridae. In: Ziegler J (Ed.) Diptera Stelviana - A dipterological perspective on a changing alpine landscape - Results from a survey of the biodiversity of Diptera (Insecta) in the Stilfserjoch National Park (Italy). Studia Dipterologica Supplement. 16.1.
- A review of the South Pacific Manota Williston (Diptera, Mycetophilidae), with the description of thirteen new species.Zootaxa4020(2):257‑288. https://doi.org/10.11646/zootaxa.4020.2.3
- Two peculiar new Orfelia Costa species from Georgia (Diptera: Keroplatidae).Entomologica Fennica24:21‑27.
- Two new Neuratelia Rondani (Diptera, Mycetophilidae) species from Western Palaearctic: a case of limited congruence between morphology and DNA sequence data.ZooKeys496:105‑129. https://doi.org/10.3897/zookeys.496.9315
- Baeopterogyna mihalyii Matile (Diptera, Mycetophilidae): association of sexes using morphological and molecular approaches with the first description of females.ZooKeys114:15‑27. https://doi.org/10.3897/zookeys.114.1364
- Three new Scandinavian species of Culicoides (Culicoides): C. boyi sp. nov., C. selandicus sp. nov. and C. kalix sp. nov. (Diptera: Ceratopogonidae).Biodiversity Data Journal3:e5823. https://doi.org/10.3897/bdj.3.e5823
- Beitrag zur Kenntnis der Ceroplatinen-Fauna Japans (Diptera, Fungivoridae).Insecta Matsumurana13:17‑32.
- New rearing records of forest-dwelling Diptera.International Journal of Dipterological Research10:133‑146.
- Fungus-gnats (Diptera, Mycetophiloidea) of Siberia.Izdatel'stvo Tomskogo Universiteta,Tomsk,308pp. [InRussian].
- Systema Dipterorum, Version 1.5. Last updated: 13 June 2013. http://www.diptera.org. Accessed on: 2016-10-15.
- Diptera diversity. Status, challenges and tools.Brill,460pp.
- Revision der europäischen Arten der Pilzmückengattung Phronia (Diptera: Mycetophilidae).Deutsch. Entomol. Zeitschr. N.F.24:305‑344. https://doi.org/10.1002/mmnd.19770240406
- Neue Pilzmückenfänge aus dem Allgäu (Diptera, Mycetophilidae).Mitteilungen der Münchner Entomologischen Gesellschaft70:15‑34.
- Fungus gnats (Diptera: Bolitophilidae, Ditomyiidae, Keroplatidae, Diadocidiidae, Mycetophilidae) in Karelia.Karelian Research Centre, Russian Academy of Sciences,Petrozavodsk,84pp. [InRussian].
- New and little known species of the fungus gnat subfamilies Mycomyinae and Sciophilinae (Diptera, Mycetophilidae) from Eastern Fennoscandia.Entomological Review81(2):207‑214.
- Fungus gnats (Diptera: Bolitophilidae, Keroplatidae, Mycetophilidae) of Pasvik strict nature reserve.Trudy Karel'skogo NC RAN. Seriya biogeografiya10:95‑104. [InRussian].
- BARCODING: bold: The Barcode of Life Data System (http://www.barcodinglife.org).Molecular Ecology Notes7(3):355‑364. https://doi.org/10.1111/j.1471-8286.2007.01678.x
- A new Neoplatyura Malloch from Finland (Diptera, Keroplatidae).Biodiversity Data Journal2:e1323. https://doi.org/10.3897/bdj.2.e1323
- New and poorly known Holarctic species of Boletina Staeger, 1840 (Diptera, Mycetophilidae).Biodiversity data journal4:e7218. https://doi.org/10.3897/BDJ.4.e7218
- Japanese Mycetophilidae (Diptera). 7. Genus Boletina Staeger.The Scientific reports of Kyoto Prefectural University (Agric.)26(197):44‑66.
- Two new species and other new records of fungus gnats (Diptera: Mycetophilidae and Keroplatidae) from Slovakia and the Czech Republic.Čas. Slez. Muz. Opava (A)58:55‑60.
- A new species of Docosia Winnertz from Central Europe, with DNA barcoding based on four gene markers (Diptera, Mycetophilidae).ZooKeys549:127‑143. https://doi.org/10.3897/zookeys.549.6925
- Molecular phylogeny of the megadiverse insect infraorder Bibionomorpha sensu lato (Diptera).PeerJ4:e2563. https://doi.org/10.7717/peerj.2563
- The adult morphology of Mycetophilidae, with a tentative phylogeny of the family (Diptera, Sciaroidea).Entomologica scandinavica Supplement50:5‑55.
- Søli GE, Vockeroth JR, Matile L (2000) Families of Sciaroidea. In: Papp L, Darvas B (Eds) Contributions to a Manual of Palaearctic Diptera (with special reference to flies of economic importance). Appendix.Science Herald
- When DNA barcoding and morphology mesh: Ceratopogonidae diversity in Finnmark, Norway.ZooKeys463:95‑131. https://doi.org/10.3897/zookeys.463.7964
- Episodic radiations in the fly tree of life.Proceedings of the National Academy of Sciences of the United States of America108(14):5690‑5695. https://doi.org/10.1073/pnas.1012675108
- Beitrag zu einer Monographie der Pilzmücken.Verh. Zool.-Bot. Ges. Wien13:637‑964.
- Holarctic fungus gnats of the genus Sciophila Meig.Nauka,Moscow,75pp. [InRussian].
- Fungus gnats of the fauna of Russia and adjacent regions. Part 1.Nauka,Moscow,288pp. [InRussian].
- Fungus gnats (Diptera, Sciaroidea) of the fauna of Russia and adjacent regions. Part 2.International Journal of Dipterological Research14:77‑386.
- New data on the fungus gnats from the Russian Far East (Diptera: Sciaroidea).Beiträge zur Entomologie46:159‑167.