|
Biodiversity Data Journal :
Taxonomic paper
|
|
Corresponding author:
Academic editor: Pierfilippo Cerretti
Received: 26 Jan 2015 | Accepted: 20 Mar 2015 | Published: 25 Mar 2015
© 2015 AJ Fleming, D. Monty Wood, Daniel Janzen, Winnie Hallwachs, M. Alex Smith
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Fleming A, Wood D, Janzen D, Hallwachs W, Smith MA (2015) Seven new species of Spathidexia Townsend (Diptera: Tachinidae) reared from caterpillars in Area de Conservación Guanacaste, Costa Rica. Biodiversity Data Journal 3: e4597. https://doi.org/10.3897/BDJ.3.e4597
|
|
Abstract
We describe seven new species of Spathidexia (Diptera: Tachinidae) reared from Area de Conservación Guanacaste (ACG), northwestern Costa Rica. All were reared from various species of ACG caterpillars during an ongoing inventory of caterpillars, their food plants and their parasitoids. By coupling morphology, photographic documentation, life history and molecular data, we provide a clear and concise description of each species. All are known to be previously undescribed as a result of a comprehensive study of the genus by DMW. Spathidexia atripalpus sp. n., Spathidexia juanvialesi sp. n., Spathidexia marioburgosi sp. n., Spathidexia luisrobertogallegosi sp. n., Spathidexia luteola sp. n., Spathidexia hernanrodriguezi sp. n. and Spathidexia aurantiaca sp. n. are all authored and described by Fleming and Wood. Minthodexiopsis Townsend is proposed by Wood as a new synonym of Spathidexia. A new combination proposed by Wood as a result of the new synonymy is S. flavicornis (Brauer & Bergenstamm) comb. n.
Keywords
Spathidexia, Diptera, Tachinidae, tropical rain forest, tropical dry forest, parasitoid fly, host-specificity, caterpillars.
Introduction
The tachinid genus Spathidexia
This work aims to build on the existing knowledge and describes seven new species of Spathidexia, all reared from caterpillars collected in ACG. In fact, all species of Spathidexia reared in ACG have been found to be undescribed species. These species are recognized based on differences in external morphology, and COI (coxI or cytochrome oxidase I) gene sequences (a.k.a. “DNA barcodes”). By coupling COI data with morphological descriptions we are able to show that abdominal markings are not only different between males and females but are also consistent within a species. As such, they are ideal tools for visual species differentiation.
The treatments reported here is focused on placing names on the species reared from ACG, thereby preparing them for later detailed ecological and behavioral accounts and studies that will normally extend across ACG ecological groups, whole ecosystems, and taxonomic assemblages much larger than a genus. However, all ACG Spathidexia reared to date in ACG are parasitoids of monocot-eating caterpillars in the Nymphalidae (Satyrinae) or Hesperiidae (Hesperiinae).
Materials and methods
Acronyms for depositories
BMNH The Natural History Museum, London, United Kingdom
CNC Canadian National Collection of Insects, Arachnids and Nematodes, Ottawa, Canada
USNM National Museum of Natural History, Washington, D.C., USA
INBio Instituto Nacional de Biodiversidad, Santo Domingo de Heredia, Costa Rica
NHMW Naturhistorisches Museum, Wien, Austria
MZUT Musei di Zoologia, Università di Torino, Torino, Italia
CAS California Academy of Sciences, San Francisco, CA, USA
Geographic area of the study and rearing intensity
All flies and rearing information described here were found by the 35+ year-old ongoing inventory of the caterpillars, their food plants and their parasitoids of the dry forest, rain forest, cloud forest, and intergrades, in the 125,000+ ha terrestrial portion of Area de Conservación Guanacaste (ACG) in northwestern Costa Rica (
In brief, caterpillars at all instars (and sometimes pupae) are found in the wild by a wide variety of search methods, and reared in captivity on the food plant species on which they were found, until they produce an adult, a parasitoid, or die of other causes. Each caterpillar is documented as an individual, as are the adult parasitic flies.
This inventory has reared about 600,000 wild-caught caterpillars since 1978. All frequencies of parasitism reported here need to be considered against this background inventory. Equally, it is patently obvious that the inventory searches some kinds of vegetation and height off the ground much more thoroughly than others, and it also searches throughout the year. Comparison of reared species of parasitoids with those collected by net or Malaise traps demonstrates that, to date, the caterpillar inventory has encountered well less than half the species of caterpillar parasitoids present in ACG. The largest unsampled void is the upper foliage of the canopy above about 3–4 m above the ground.
Imaging and dissections
Our descriptions of new species are deliberately brief and only include some differentiating descriptions of body parts and colors that are commonly used in tachinid identification. These brief descriptions are complemented with an extensive series of color photos of every species to illustrate the readily-observed differences among these species.
Habitus photographs were taken using a Canon T3i digital SLR, using a 65mm Macro Photo Lens 1:2.8 (MP–E 65mm), mounted on a microscope track stand (AmScope, Model: TS200) modified to accept a Manfrotto QR 200PL–14 quick release plate. Images were shot in aperture priority, allowing the camera to control shutter speed at f/4.5. Stacks of 40 images were taken at equal distance increments. Illumination was provided with a homemade reflective dome (instruction for dome creation can be found at: http://www.cdfa.ca.gov/plant/ppd/entomology/Dome/kd–200.html) placed over a 144 LED ringlight (AmScope, Model: LED–144–YK).
The photographic series were processed using Photoshop CS6, and digitally stacked with Zerene Stacker Software v1.04. This produced a final composite image maximizing image quality and depth of field.
The terminology used for components of the terminalia (which refers here only to the sclerotized parts of the genitalia, and not to the soft internal structures) and other body parts, follows
All specimens listed as examined are considered paratypes, except for the holotype which is noted separately.
Wherever a specimen label has been cited, the information is presented using the following symbols: /, indicates the end of a line; //, indicates the end of a label. Labels are presented from top (closest to the specimen) to bottom, with any comments about the label being given in square brackets.
Voucher specimen management
All caterpillars reared from the ACG efforts receive a unique voucher code in the format of yy–SRNP–xxxxx. Any parasitoid emerging from this caterpillar receives the same voucher code, and then if/when later the parasitoid is dealt with individually, it receives a second voucher code unique to it, in the format of DHJPARxxxxxxx. The voucher codes and collateral data assigned to both host and emergent parasitoids are available at http://janzen.bio.upenn.edu/caterpillars/database.lasso. To date, all DHJPARxxxxxxx coded tachinids have had one leg removed for attempted DNA barcoding at the Biodiversity Institute of Ontario (BIO) at the University of Guelph, with all collateral data and all successful barcodes permanently and publically deposited in the Barcode of Life Data System (BOLD, www.boldsystems.org) (
Inventoried Tachinidae were collected under Costa Rican government research permits issued to DHJ since 1978, and likewise exported under permit by DHJ from Costa Rica to Philadelphia, and then to the final depository in the Canadian National Insect collection in Ottawa, Canada. Tachinid identifications for the inventory were done by DHJ in coordination with a) visual inspection by AJF and DMW, b) DNA barcoding by BIO, MAS, and BOLD, and c) correlation with host caterpillar identifications by DHJ and WH through the inventory itself. Dates of capture of each reared fly in the inventory are the dates of eclosion of the fly, and not the date of capture of the caterpillar. This is because the fly eclosion date is much more representative of the time when that fly species is on the wing than is the time of capture of the caterpillar or (rarely) finding a parasitized pupa. However, the collector listed is the parataxonomist who found the caterpillar, rather than the person who retrieved the newly eclosed fly from its rearing bag or bottle, and processed it by freezing, pinning, labeling and oven–drying. Fly biology and degrees of parasitization by these flies will be the detailed subject of later papers.
DNA barcoding
DNA barcodes (standardised 5’ region of the mitochondrial cytochrome c oxidase I (COI) gene) for all ACG inventory specimens were obtained using DNA extracts prepared from single legs using a glass fibre protocol (
Generic synonyms of Spathidexia
Spathidexia
Gymnopalpus
Minthodexiopsis
Minthohoughia
Stenaulacodoria
Previously described species included in Spathidexia
antillensis
brasiliensis
cerussata
cinereicollis
clemonsi
creolensis
cylindrica
dicta
dunningii
rasilis
elegans
flavicornis
nexa
niveomarginata
pallida
reinhardi
setipennis
spatulata
*The new combination is proposed by DMW and result from new generic synonymy and the examination of the type material of other nominal species. Type label information is included for those species.
Diagnosis of the genus Spathidexia
Taxon treatments
Spathidexia atripalpus , sp. n.
-
scientificName: Spathidexia atripalpus; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:atripalpus; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Ingas; verbatimElevation:580; verbatimLatitude:11.00311; verbatimLongitude:-85.288; verbatimCoordinateSystem:Decimal; decimalLatitude:11.00311; decimalLongitude:-85.288; samplingProtocol:reared from caterpillar of Magneuptychia libye (Nymphalidae); verbatimEventDate:19/Mar/12; individualID:DHJPAR0048611; individualCount:1; sex:M; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0048611; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:12-SRNP-30227; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia atripalpus; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:atripalpus; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector El Hacha; verbatimLocality:Estacion Los Almendros; verbatimElevation:290; verbatimLatitude:11.03226; verbatimLongitude:-85.52776; verbatimCoordinateSystem:Decimal; decimalLatitude:11.03226; decimalLongitude:-85.52776; samplingProtocol:reared from caterpillar of Pierella pallida (Nymphalidae); verbatimEventDate:24/Jul/99; individualID:DHJPAR0018619; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0018619; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:99-SRNP-2997; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia atripalpus; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:atripalpus; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Ingas; verbatimElevation:580; verbatimLatitude:11.00311; verbatimLongitude:-85.42041; verbatimCoordinateSystem:Decimal; decimalLatitude:11.00311; decimalLongitude:-85.42041; samplingProtocol:reared from caterpillar of Pareuptychia ocirrhoe (Nymphalidae); verbatimEventDate:26/Jan/12; individualID:DHJPAR0048578; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0048578; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:12-SRNP-33263; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
Description
Male (Fig.






Female (Fig.
Diagnosis
The presence of black palpi is its strongest diagnostic character, differentiating it from almost all other species. This species differs from S. atypica, which also has black(ish) palpi, in having 3 postsutural dorsocentral bristles, bare eyes, and the presence of small black hairs covering entire body especially prominent extending from anepimeron and underside of T1+2. Differs, from S. antillensis, which also bears two bold thoracic vittae but does not bear black palpi.
Etymology
From the Latin “ater”, color black, and the noun “palpus”, literally the palm of the hand, in reference to the black palpi present in both sexes of this species.
Distribution
Costa Rica, ACG, Prov. Guanacaste, rain forest and rain forest/dry forest ecotone, 290-580m elevation.
Ecology
Reared from four species of grass-eating rain forest satyrine Nymphalidae (4 records). One larva per host.
Spathidexia marioburgosi , sp. n.
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Bosque Aguirre; verbatimElevation:620; verbatimLatitude:11.001; verbatimLongitude:-85.438; verbatimCoordinateSystem:Decimal; decimalLatitude:11.001; decimalLongitude:-85.438; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:14-Sep-2008; individualID:DHJPAR0029985; individualCount:1; sex:M; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0029985; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-22719; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Rio Areno; verbatimElevation:460; verbatimLatitude:10.914; verbatimLongitude:-85.382; verbatimCoordinateSystem:Decimal; decimalLatitude:10.914; decimalLongitude:-85.382; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:07-Nov-2005; individualID:DHJPAR0005445; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0005445; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-6295; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Sendero Huerta; verbatimElevation:527; verbatimLatitude:10.931; verbatimLongitude:-85.372; verbatimCoordinateSystem:Decimal; decimalLatitude:10.931; decimalLongitude:-85.372; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:01-Dec-2005; individualID:DHJPAR0005453; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0005453; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-6890; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Rio Blanco Abajo; verbatimElevation:500; verbatimLatitude:10.9; verbatimLongitude:-85.373; verbatimCoordinateSystem:Decimal; decimalLatitude:10.9; decimalLongitude:-85.373; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:01-Dec-2005; individualID:DHJPAR0005454; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0005454; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-6517; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Sendero Huerta; verbatimElevation:527; verbatimLatitude:10.931; verbatimLongitude:-85.372; verbatimCoordinateSystem:Decimal; decimalLatitude:10.931; decimalLongitude:-85.372; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:13-Dec-2005; individualID:DHJPAR0005458; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0005458; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-7188; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Vado Rio Cucaracho; verbatimElevation:640; verbatimLatitude:10.87; verbatimLongitude:-85.392; verbatimCoordinateSystem:Decimal; decimalLatitude:10.87; decimalLongitude:-85.392; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:09-Dec-2005; individualID:DHJPAR0005459; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0005459; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-7049; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Montanya Figueres; verbatimElevation:460; verbatimLatitude:10.884; verbatimLongitude:-85.291; verbatimCoordinateSystem:Decimal; decimalLatitude:10.884; decimalLongitude:-85.291; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:09-Nov-2005; individualID:DHJPAR0005463; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0005463; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-42898; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Estacion Caribe; verbatimElevation:415; verbatimLatitude:10.902; verbatimLongitude:-85.275; verbatimCoordinateSystem:Decimal; decimalLatitude:10.902; decimalLongitude:-85.275; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:03-Dec-2005; individualID:DHJPAR0005467; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0005467; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-43349; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Sendero Juntas; verbatimElevation:400; verbatimLatitude:10.907; verbatimLongitude:-85.288; verbatimCoordinateSystem:Decimal; decimalLatitude:10.907; decimalLongitude:-85.288; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:20-Nov-2005; individualID:DHJPAR0005470; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0005470; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-43130; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Sendero Guaca; verbatimElevation:400; verbatimLatitude:10.906; verbatimLongitude:-85.283; verbatimCoordinateSystem:Decimal; decimalLatitude:10.906; decimalLongitude:-85.283; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:25-Dec-2005; individualID:DHJPAR0005471; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0005471; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-43431; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Sendero Puertas; verbatimElevation:400; verbatimLatitude:11.011; verbatimLongitude:-85.488; verbatimCoordinateSystem:Decimal; decimalLatitude:11.011; decimalLongitude:-85.488; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:08-Dec-2005; individualID:DHJPAR0005473; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0005473; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-25074; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Finca San Gabriel; verbatimElevation:645; verbatimLatitude:10.878; verbatimLongitude:-85.393; verbatimCoordinateSystem:Decimal; decimalLatitude:10.878; decimalLongitude:-85.393; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:06-Feb-2006; individualID:DHJPAR0011559; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0011559; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-8130; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Estacion Caribe; verbatimElevation:415; verbatimLatitude:10.902; verbatimLongitude:-85.275; verbatimCoordinateSystem:Decimal; decimalLatitude:10.902; decimalLongitude:-85.275; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:03-Dec-2005; individualID:DHJPAR0011572; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0011572; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-43349; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Uncaria; verbatimElevation:370; verbatimLatitude:11.018; verbatimLongitude:-85.474; verbatimCoordinateSystem:Decimal; decimalLatitude:11.018; decimalLongitude:-85.474; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:06-Oct-2003; individualID:DHJPAR0015271; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0015271; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-28308; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Rio Blanco Abajo; verbatimElevation:500; verbatimLatitude:10.9; verbatimLongitude:-85.373; verbatimCoordinateSystem:Decimal; decimalLatitude:10.9; decimalLongitude:-85.373; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:16-Jul-2006; individualID:DHJPAR0016073; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0016073; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-4769; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Mundo Nuevo; verbatimLocality:Quebrada Tibio Perla; verbatimElevation:330; verbatimLatitude:10.763; verbatimLongitude:-85.43; verbatimCoordinateSystem:Decimal; decimalLatitude:10.763; decimalLongitude:-85.43; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:21-Sep-2006; individualID:DHJPAR0016107; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0016107; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-58077; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Finca San Gabriel; verbatimElevation:645; verbatimLatitude:10.878; verbatimLongitude:-85.393; verbatimCoordinateSystem:Decimal; decimalLatitude:10.878; decimalLongitude:-85.393; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:12-Oct-2006; individualID:DHJPAR0016178; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0016178; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-7631; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Mundo Nuevo; verbatimLocality:Vado Huacas; verbatimElevation:490; verbatimLatitude:10.755; verbatimLongitude:-85.391; verbatimCoordinateSystem:Decimal; decimalLatitude:10.755; decimalLongitude:-85.391; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:11-Oct-2006; individualID:DHJPAR0016180; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0016180; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-58374; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Mundo Nuevo; verbatimLocality:Vado Huacas; verbatimElevation:490; verbatimLatitude:10.755; verbatimLongitude:-85.391; verbatimCoordinateSystem:Decimal; decimalLatitude:10.755; decimalLongitude:-85.391; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:11-Oct-2006; individualID:DHJPAR0016236; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0016236; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-58370; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Finca San Gabriel; verbatimElevation:645; verbatimLatitude:10.878; verbatimLongitude:-85.393; verbatimCoordinateSystem:Decimal; decimalLatitude:10.878; decimalLongitude:-85.393; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:01-Sep-2006; individualID:DHJPAR0016242; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0016242; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-6683; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Puente Palma; verbatimElevation:460; verbatimLatitude:10.916; verbatimLongitude:-85.379; verbatimCoordinateSystem:Decimal; decimalLatitude:10.916; decimalLongitude:-85.379; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:08-Sep-2006; individualID:DHJPAR0016313; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0016313; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-6796; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Gongora Bananal; verbatimElevation:600; verbatimLatitude:10.889; verbatimLongitude:-85.476; verbatimCoordinateSystem:Decimal; decimalLatitude:10.889; decimalLongitude:-85.476; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:05-Sep-2006; individualID:DHJPAR0016330; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0016330; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-46866; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Puente Gongora; verbatimElevation:540; verbatimLatitude:10.885; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.885; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:24-Sep-2006; individualID:DHJPAR0016488; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0016488; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-47185; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Sendero Llano; verbatimElevation:400; verbatimLatitude:10.903; verbatimLongitude:-85.29; verbatimCoordinateSystem:Decimal; decimalLatitude:10.903; decimalLongitude:-85.29; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:06-Jan-2007; individualID:DHJPAR0016537; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0016537; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-44678; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Puente Gongora; verbatimElevation:540; verbatimLatitude:10.885; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.885; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:12-Dec-2006; individualID:DHJPAR0016600; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0016600; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-48089; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Puente Palma; verbatimElevation:460; verbatimLatitude:10.916; verbatimLongitude:-85.379; verbatimCoordinateSystem:Decimal; decimalLatitude:10.916; decimalLongitude:-85.379; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:31-Oct-2006; individualID:DHJPAR0019550; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019550; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-8343; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:El Ensayo; verbatimLocality:Metereologico; verbatimElevation:330; verbatimLatitude:10.76261; verbatimLongitude:-85.42979; verbatimCoordinateSystem:Decimal; decimalLatitude:10.76261; decimalLongitude:-85.42979; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:18-May-2007; individualID:DHJPAR0019612; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019612; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-65140; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Mundo Nuevo; verbatimLocality:Quebrada Tibio Perla; verbatimElevation:330; verbatimLatitude:10.763; verbatimLongitude:-85.43; verbatimCoordinateSystem:Decimal; decimalLatitude:10.763; decimalLongitude:-85.43; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:10-Jun-2007; individualID:DHJPAR0019613; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019613; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-56562; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Mundo Nuevo; verbatimLocality:Quebrada Tibio Perla; verbatimElevation:330; verbatimLatitude:10.763; verbatimLongitude:-85.43; verbatimCoordinateSystem:Decimal; decimalLatitude:10.763; decimalLongitude:-85.43; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:10-Jun-2007; individualID:DHJPAR0019709; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019709; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-56533; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Finca San Gabriel; verbatimElevation:645; verbatimLatitude:10.878; verbatimLongitude:-85.393; verbatimCoordinateSystem:Decimal; decimalLatitude:10.878; decimalLongitude:-85.393; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:17-May-2007; individualID:DHJPAR0019710; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019710; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-1851; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Finca San Gabriel; verbatimElevation:645; verbatimLatitude:10.878; verbatimLongitude:-85.393; verbatimCoordinateSystem:Decimal; decimalLatitude:10.878; decimalLongitude:-85.393; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:18-May-2007; individualID:DHJPAR0019711; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019711; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-1852; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Finca San Gabriel; verbatimElevation:645; verbatimLatitude:10.878; verbatimLongitude:-85.393; verbatimCoordinateSystem:Decimal; decimalLatitude:10.878; decimalLongitude:-85.393; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:18-May-2007; individualID:DHJPAR0019801; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019801; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-1852; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Mundo Nuevo; verbatimLocality:Quebrada Tibio Perla; verbatimElevation:330; verbatimLatitude:10.763; verbatimLongitude:-85.43; verbatimCoordinateSystem:Decimal; decimalLatitude:10.763; decimalLongitude:-85.43; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:05-Jun-2007; individualID:DHJPAR0019954; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019954; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-56555; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Mundo Nuevo; verbatimLocality:Vado Miramonte; verbatimElevation:305; verbatimLatitude:10.772; verbatimLongitude:-85.434; verbatimCoordinateSystem:Decimal; decimalLatitude:10.772; decimalLongitude:-85.434; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:17-Aug-2007; individualID:DHJPAR0020920; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0020920; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-58809; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Mundo Nuevo; verbatimLocality:Quebrada Tibio Perla; verbatimElevation:330; verbatimLatitude:10.763; verbatimLongitude:-85.43; verbatimCoordinateSystem:Decimal; decimalLatitude:10.763; decimalLongitude:-85.43; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:31-Aug-2007; individualID:DHJPAR0020936; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0020936; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-58951; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Mundo Nuevo; verbatimLocality:Quebrada Tibio Perla; verbatimElevation:330; verbatimLatitude:10.763; verbatimLongitude:-85.43; verbatimCoordinateSystem:Decimal; decimalLatitude:10.763; decimalLongitude:-85.43; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:25-Jul-2007; individualID:DHJPAR0020937; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0020937; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-58016; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Mundo Nuevo; verbatimLocality:Vado Miramonte; verbatimElevation:305; verbatimLatitude:10.772; verbatimLongitude:-85.434; verbatimCoordinateSystem:Decimal; decimalLatitude:10.772; decimalLongitude:-85.434; samplingProtocol:reared from caterpillar of Talides Burns02 (Hesperiidae); verbatimEventDate:09-Aug-2007; individualID:DHJPAR0020938; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0020938; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-58356; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Mundo Nuevo; verbatimLocality:Vado Miramonte; verbatimElevation:305; verbatimLatitude:10.772; verbatimLongitude:-85.434; verbatimCoordinateSystem:Decimal; decimalLatitude:10.772; decimalLongitude:-85.434; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:07-Aug-2007; individualID:DHJPAR0021856; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0021856; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-58377; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Mundo Nuevo; verbatimLocality:Sendero Aguacate; verbatimElevation:335; verbatimLatitude:10.769; verbatimLongitude:-85.435; verbatimCoordinateSystem:Decimal; decimalLatitude:10.769; decimalLongitude:-85.435; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:03-Sep-2007; individualID:DHJPAR0022284; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022284; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-59237; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Mundo Nuevo; verbatimLocality:Vado Miramonte; verbatimElevation:305; verbatimLatitude:10.772; verbatimLongitude:-85.434; verbatimCoordinateSystem:Decimal; decimalLatitude:10.772; decimalLongitude:-85.434; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:17-Aug-2007; individualID:DHJPAR0022859; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022859; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-58809; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Quebrada Cementerio; verbatimElevation:700; verbatimLatitude:10.871; verbatimLongitude:-85.387; verbatimCoordinateSystem:Decimal; decimalLatitude:10.871; decimalLongitude:-85.387; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:19-Sep-2000; individualID:DHJPAR0022860; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022860; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:00-SRNP-12307; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Quebrada Cementerio; verbatimElevation:700; verbatimLatitude:10.871; verbatimLongitude:-85.387; verbatimCoordinateSystem:Decimal; decimalLatitude:10.871; decimalLongitude:-85.387; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:15-Sep-2000; individualID:DHJPAR0022861; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022861; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:00-SRNP-12306; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Quebrada Cementerio; verbatimElevation:700; verbatimLatitude:10.871; verbatimLongitude:-85.387; verbatimCoordinateSystem:Decimal; decimalLatitude:10.871; decimalLongitude:-85.387; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:14-Sep-2000; individualID:DHJPAR0022862; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022862; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:00-SRNP-12303; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Camino Rio Francia; verbatimElevation:410; verbatimLatitude:10.904; verbatimLongitude:-85.287; verbatimCoordinateSystem:Decimal; decimalLatitude:10.904; decimalLongitude:-85.287; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:06-Dec-2000; individualID:DHJPAR0022863; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022863; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:00-SRNP-20936; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Camino Rio Francia; verbatimElevation:410; verbatimLatitude:10.904; verbatimLongitude:-85.287; verbatimCoordinateSystem:Decimal; decimalLatitude:10.904; decimalLongitude:-85.287; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:12-Dec-2000; individualID:DHJPAR0022864; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022864; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:00-SRNP-20919; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Toma Agua; verbatimElevation:1140; verbatimLatitude:10.928; verbatimLongitude:-85.467; verbatimCoordinateSystem:Decimal; decimalLatitude:10.928; decimalLongitude:-85.467; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:23-Nov-2000; individualID:DHJPAR0022865; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022865; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:00-SRNP-10863; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Quebrada Cementerio; verbatimElevation:700; verbatimLatitude:10.871; verbatimLongitude:-85.387; verbatimCoordinateSystem:Decimal; decimalLatitude:10.871; decimalLongitude:-85.387; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:30-Nov-2000; individualID:DHJPAR0022866; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022866; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:00-SRNP-21776; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Cuesta Caimito; verbatimElevation:640; verbatimLatitude:10.891; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.891; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:09-Sep-2000; individualID:DHJPAR0022867; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022867; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:00-SRNP-10300; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Quebrada Serrano; verbatimElevation:585; verbatimLatitude:11; verbatimLongitude:-85.456; verbatimCoordinateSystem:Decimal; decimalLatitude:11; decimalLongitude:-85.456; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:05-Nov-1999; individualID:DHJPAR0022868; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022868; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:99-SRNP-15104; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Gongora Bananal; verbatimElevation:600; verbatimLatitude:10.889; verbatimLongitude:-85.476; verbatimCoordinateSystem:Decimal; decimalLatitude:10.889; decimalLongitude:-85.476; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:22-Sep-2004; individualID:DHJPAR0022869; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022869; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-48259; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Quebrada Heliconia; verbatimElevation:390; verbatimLatitude:10.886; verbatimLongitude:-85.492; verbatimCoordinateSystem:Decimal; decimalLatitude:10.886; decimalLongitude:-85.492; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:30-Sep-2004; individualID:DHJPAR0022870; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022870; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-48421; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Gongora Bananal; verbatimElevation:600; verbatimLatitude:10.889; verbatimLongitude:-85.476; verbatimCoordinateSystem:Decimal; decimalLatitude:10.889; decimalLongitude:-85.476; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:23-Sep-2004; individualID:DHJPAR0022871; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022871; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-48245; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Rio Blanco Abajo; verbatimElevation:500; verbatimLatitude:10.9; verbatimLongitude:-85.373; verbatimCoordinateSystem:Decimal; decimalLatitude:10.9; decimalLongitude:-85.373; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:04-Aug-2000; individualID:DHJPAR0022872; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022872; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:00-SRNP-11612; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Cuesta Caimito; verbatimElevation:640; verbatimLatitude:10.891; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.891; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:10-Aug-2004; individualID:DHJPAR0022873; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022873; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-47061; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Cuesta Caimito; verbatimElevation:640; verbatimLatitude:10.891; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.891; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:09-Aug-2004; individualID:DHJPAR0022874; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022874; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-47065; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Camino Mangos; verbatimElevation:480; verbatimLatitude:11.008; verbatimLongitude:-85.479; verbatimCoordinateSystem:Decimal; decimalLatitude:11.008; decimalLongitude:-85.479; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:05-Sep-2003; individualID:DHJPAR0022875; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022875; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-18879; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Uncaria; verbatimElevation:370; verbatimLatitude:11.018; verbatimLongitude:-85.474; verbatimCoordinateSystem:Decimal; decimalLatitude:11.018; decimalLongitude:-85.474; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:06-Oct-2003; individualID:DHJPAR0022876; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022876; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-28308; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Camino Porvenir; verbatimElevation:383; verbatimLatitude:10.904; verbatimLongitude:-85.26; verbatimCoordinateSystem:Decimal; decimalLatitude:10.904; decimalLongitude:-85.26; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:08-Oct-2003; individualID:DHJPAR0022877; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022877; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-12458.1; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Quebrada Trigal; verbatimElevation:290; verbatimLatitude:11.027; verbatimLongitude:-85.495; verbatimCoordinateSystem:Decimal; decimalLatitude:11.027; decimalLongitude:-85.495; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:15-Sep-2003; individualID:DHJPAR0022878; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022878; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-19475; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Quebrada Trigal; verbatimElevation:290; verbatimLatitude:11.027; verbatimLongitude:-85.495; verbatimCoordinateSystem:Decimal; decimalLatitude:11.027; decimalLongitude:-85.495; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:12-Sep-2003; individualID:DHJPAR0022879; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022879; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-19466; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Rio Blanco Abajo; verbatimElevation:500; verbatimLatitude:10.9; verbatimLongitude:-85.373; verbatimCoordinateSystem:Decimal; decimalLatitude:10.9; decimalLongitude:-85.373; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:22-Sep-2003; individualID:DHJPAR0022880; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022880; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-7910; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Quebrada Trigal; verbatimElevation:290; verbatimLatitude:11.027; verbatimLongitude:-85.495; verbatimCoordinateSystem:Decimal; decimalLatitude:11.027; decimalLongitude:-85.495; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:05-Sep-2003; individualID:DHJPAR0022881; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022881; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-19471; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Cuesta Caimito; verbatimElevation:640; verbatimLatitude:10.891; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.891; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:09-Aug-2004; individualID:DHJPAR0022882; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022882; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-47065; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Cementerio Viejo; verbatimElevation:570; verbatimLatitude:10.881; verbatimLongitude:-85.389; verbatimCoordinateSystem:Decimal; decimalLatitude:10.881; decimalLongitude:-85.389; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:11-May-2004; individualID:DHJPAR0022883; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022883; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-1867; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Quebrada Suamposa; verbatimElevation:290; verbatimLatitude:11.024; verbatimLongitude:-85.491; verbatimCoordinateSystem:Decimal; decimalLatitude:11.024; decimalLongitude:-85.491; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:04-Jul-2004; individualID:DHJPAR0022884; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022884; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-22687; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Rio Blanco Abajo; verbatimElevation:500; verbatimLatitude:10.9; verbatimLongitude:-85.373; verbatimCoordinateSystem:Decimal; decimalLatitude:10.9; decimalLongitude:-85.373; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:04-Nov-2003; individualID:DHJPAR0022885; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022885; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-8968; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Finca San Gabriel; verbatimElevation:645; verbatimLatitude:10.878; verbatimLongitude:-85.393; verbatimCoordinateSystem:Decimal; decimalLatitude:10.878; decimalLongitude:-85.393; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:15-Feb-2003; individualID:DHJPAR0022886; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022886; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-95; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Quebrada Trigal; verbatimElevation:290; verbatimLatitude:11.027; verbatimLongitude:-85.495; verbatimCoordinateSystem:Decimal; decimalLatitude:11.027; decimalLongitude:-85.495; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:30-Oct-2003; individualID:DHJPAR0022887; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022887; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-29175; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Margarita; verbatimElevation:380; verbatimLatitude:11.032; verbatimLongitude:-85.44; verbatimCoordinateSystem:Decimal; decimalLatitude:11.032; decimalLongitude:-85.44; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:07-Nov-2003; individualID:DHJPAR0022888; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022888; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-29689; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Sendero Puertas; verbatimElevation:400; verbatimLatitude:11.011; verbatimLongitude:-85.488; verbatimCoordinateSystem:Decimal; decimalLatitude:11.011; decimalLongitude:-85.488; samplingProtocol:reared from caterpillar of Talides Burns02 (Hesperiidae); verbatimEventDate:06-Nov-2003; individualID:DHJPAR0022889; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022889; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-29255; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Pasmompa; verbatimElevation:440; verbatimLatitude:11.019; verbatimLongitude:-85.41; verbatimCoordinateSystem:Decimal; decimalLatitude:11.019; decimalLongitude:-85.41; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:05-Feb-2004; individualID:DHJPAR0022890; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022890; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-30107; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Quebrada Lajosa; verbatimElevation:400; verbatimLatitude:11.033; verbatimLongitude:-85.429; verbatimCoordinateSystem:Decimal; decimalLatitude:11.033; decimalLongitude:-85.429; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:31-Jan-2004; individualID:DHJPAR0022891; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022891; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-20014; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Puente Palma; verbatimElevation:460; verbatimLatitude:10.916; verbatimLongitude:-85.379; verbatimCoordinateSystem:Decimal; decimalLatitude:10.916; decimalLongitude:-85.379; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:31-Mar-2004; individualID:DHJPAR0022892; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022892; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-1130; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Finca San Gabriel; verbatimElevation:645; verbatimLatitude:10.878; verbatimLongitude:-85.393; verbatimCoordinateSystem:Decimal; decimalLatitude:10.878; decimalLongitude:-85.393; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:12-Dec-2003; individualID:DHJPAR0022893; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022893; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-34139; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Cementerio Viejo; verbatimElevation:570; verbatimLatitude:10.881; verbatimLongitude:-85.389; verbatimCoordinateSystem:Decimal; decimalLatitude:10.881; decimalLongitude:-85.389; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:25-Jun-2004; individualID:DHJPAR0022894; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022894; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-2436; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Quebrada Suamposa; verbatimElevation:290; verbatimLatitude:11.024; verbatimLongitude:-85.491; verbatimCoordinateSystem:Decimal; decimalLatitude:11.024; decimalLongitude:-85.491; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:04-Jul-2004; individualID:DHJPAR0022895; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022895; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-22687; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Sendero Carmona; verbatimElevation:670; verbatimLatitude:10.876; verbatimLongitude:-85.386; verbatimCoordinateSystem:Decimal; decimalLatitude:10.876; decimalLongitude:-85.386; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:02-Aug-2004; individualID:DHJPAR0022896; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022896; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-3396; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Cuesta Caimito; verbatimElevation:640; verbatimLatitude:10.891; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.891; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:30-Jul-2004; individualID:DHJPAR0022897; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022897; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-47058; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Camino Rio Francia; verbatimElevation:410; verbatimLatitude:10.904; verbatimLongitude:-85.287; verbatimCoordinateSystem:Decimal; decimalLatitude:10.904; decimalLongitude:-85.287; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:25-Jan-2003; individualID:DHJPAR0022898; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022898; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-21538; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Rio Blanco Abajo; verbatimElevation:500; verbatimLatitude:10.9; verbatimLongitude:-85.373; verbatimCoordinateSystem:Decimal; decimalLatitude:10.9; decimalLongitude:-85.373; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:12-Jan-2003; individualID:DHJPAR0022899; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022899; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-20361; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Toma Agua; verbatimElevation:1140; verbatimLatitude:10.928; verbatimLongitude:-85.467; verbatimCoordinateSystem:Decimal; decimalLatitude:10.928; decimalLongitude:-85.467; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:19-Aug-2002; individualID:DHJPAR0022900; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022900; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-9934; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Guacimos; verbatimElevation:380; verbatimLatitude:11.015; verbatimLongitude:-85.475; verbatimCoordinateSystem:Decimal; decimalLatitude:11.015; decimalLongitude:-85.475; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:16-Sep-2002; individualID:DHJPAR0022901; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022901; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-28992; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector El Hacha; verbatimLocality:Finca Araya; verbatimElevation:295; verbatimLatitude:11.015; verbatimLongitude:-85.511; verbatimCoordinateSystem:Decimal; decimalLatitude:11.015; decimalLongitude:-85.511; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:09-Dec-2002; individualID:DHJPAR0022902; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022902; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-33351; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Camino Mangos; verbatimElevation:480; verbatimLatitude:11.008; verbatimLongitude:-85.479; verbatimCoordinateSystem:Decimal; decimalLatitude:11.008; decimalLongitude:-85.479; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:08-Sep-2002; individualID:DHJPAR0022903; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022903; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-28345; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Cuesta Caimito; verbatimElevation:640; verbatimLatitude:10.891; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.891; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:13-Sep-2004; individualID:DHJPAR0022904; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022904; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-47949; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Camino Mangos; verbatimElevation:480; verbatimLatitude:11.008; verbatimLongitude:-85.479; verbatimCoordinateSystem:Decimal; decimalLatitude:11.008; decimalLongitude:-85.479; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:05-Sep-2002; individualID:DHJPAR0022905; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022905; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-27830; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Camino Mangos; verbatimElevation:480; verbatimLatitude:11.008; verbatimLongitude:-85.479; verbatimCoordinateSystem:Decimal; decimalLatitude:11.008; decimalLongitude:-85.479; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:02-Sep-2002; individualID:DHJPAR0022906; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022906; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-27838; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Sendero Corredor; verbatimElevation:620; verbatimLatitude:10.879; verbatimLongitude:-85.39; verbatimCoordinateSystem:Decimal; decimalLatitude:10.879; decimalLongitude:-85.39; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:21-Jan-2004; individualID:DHJPAR0022907; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022907; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-34815; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Sendero Puertas; verbatimElevation:400; verbatimLatitude:11.011; verbatimLongitude:-85.488; verbatimCoordinateSystem:Decimal; decimalLatitude:11.011; decimalLongitude:-85.488; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:06-Nov-2003; individualID:DHJPAR0022908; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022908; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-29255; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Puente Gongora; verbatimElevation:540; verbatimLatitude:10.885; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.885; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:11-Sep-2005; individualID:DHJPAR0022909; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022909; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-47225; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Puente Gongora; verbatimElevation:540; verbatimLatitude:10.885; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.885; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:18-Sep-2005; individualID:DHJPAR0022910; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022910; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-47226; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Puente Gongora; verbatimElevation:540; verbatimLatitude:10.885; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.885; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:12-Sep-2005; individualID:DHJPAR0022911; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022911; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-47373; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Quebrada Cementerio; verbatimElevation:700; verbatimLatitude:10.871; verbatimLongitude:-85.387; verbatimCoordinateSystem:Decimal; decimalLatitude:10.871; decimalLongitude:-85.387; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:01-Sep-2005; individualID:DHJPAR0022912; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022912; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-4221; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Palmas; verbatimElevation:675; verbatimLatitude:10.896; verbatimLongitude:-85.474; verbatimCoordinateSystem:Decimal; decimalLatitude:10.896; decimalLongitude:-85.474; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:29-Aug-2005; individualID:DHJPAR0022913; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022913; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-47041; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Puente Gongora; verbatimElevation:540; verbatimLatitude:10.885; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.885; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:21-Sep-2005; individualID:DHJPAR0022914; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022914; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-47477; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Puente Gongora; verbatimElevation:540; verbatimLatitude:10.885; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.885; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:21-Sep-2005; individualID:DHJPAR0022915; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022915; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-47479; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Sendero Platanal; verbatimElevation:335; verbatimLatitude:10.849; verbatimLongitude:-85.253; verbatimCoordinateSystem:Decimal; decimalLatitude:10.849; decimalLongitude:-85.253; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:16-Jul-2003; individualID:DHJPAR0022916; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022916; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-11367; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Sendero Corredor; verbatimElevation:620; verbatimLatitude:10.879; verbatimLongitude:-85.39; verbatimCoordinateSystem:Decimal; decimalLatitude:10.879; decimalLongitude:-85.39; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:30-Oct-2002; individualID:DHJPAR0022917; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022917; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-19519; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Camino Mangos; verbatimElevation:480; verbatimLatitude:11.008; verbatimLongitude:-85.479; verbatimCoordinateSystem:Decimal; decimalLatitude:11.008; decimalLongitude:-85.479; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:16-Sep-2002; individualID:DHJPAR0022918; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022918; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-29143; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Guacimos; verbatimElevation:380; verbatimLatitude:11.015; verbatimLongitude:-85.475; verbatimCoordinateSystem:Decimal; decimalLatitude:11.015; decimalLongitude:-85.475; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:16-Sep-2002; individualID:DHJPAR0022919; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022919; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-28992; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Toma Agua; verbatimElevation:1140; verbatimLatitude:10.928; verbatimLongitude:-85.467; verbatimCoordinateSystem:Decimal; decimalLatitude:10.928; decimalLongitude:-85.467; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:24-Aug-2002; individualID:DHJPAR0022920; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022920; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-9935; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Toma Agua; verbatimElevation:1140; verbatimLatitude:10.928; verbatimLongitude:-85.467; verbatimCoordinateSystem:Decimal; decimalLatitude:10.928; decimalLongitude:-85.467; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:17-Aug-2002; individualID:DHJPAR0022921; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022921; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-9933; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Rio Blanco Abajo; verbatimElevation:500; verbatimLatitude:10.9; verbatimLongitude:-85.373; verbatimCoordinateSystem:Decimal; decimalLatitude:10.9; decimalLongitude:-85.373; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:04-Nov-2003; individualID:DHJPAR0022922; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022922; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-8968; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Puente Gongora; verbatimElevation:540; verbatimLatitude:10.885; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.885; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:30-Nov-2003; individualID:DHJPAR0022923; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022923; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-23786; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Quebrada Otilio; verbatimElevation:550; verbatimLatitude:10.89; verbatimLongitude:-85.48; verbatimCoordinateSystem:Decimal; decimalLatitude:10.89; decimalLongitude:-85.48; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:06-Sep-2004; individualID:DHJPAR0022924; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022924; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-48016; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Sendero Corredor; verbatimElevation:620; verbatimLatitude:10.879; verbatimLongitude:-85.39; verbatimCoordinateSystem:Decimal; decimalLatitude:10.879; decimalLongitude:-85.39; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:31-Dec-2001; individualID:DHJPAR0022925; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022925; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:01-SRNP-22877; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Guacimos; verbatimElevation:380; verbatimLatitude:11.015; verbatimLongitude:-85.475; verbatimCoordinateSystem:Decimal; decimalLatitude:11.015; decimalLongitude:-85.475; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:04-Sep-2002; individualID:DHJPAR0022926; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022926; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-27700; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Rio Blanco Abajo; verbatimElevation:500; verbatimLatitude:10.9; verbatimLongitude:-85.373; verbatimCoordinateSystem:Decimal; decimalLatitude:10.9; decimalLongitude:-85.373; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:24-Apr-2002; individualID:DHJPAR0022927; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022927; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-2319; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Rio Blanco Abajo; verbatimElevation:500; verbatimLatitude:10.9; verbatimLongitude:-85.373; verbatimCoordinateSystem:Decimal; decimalLatitude:10.9; decimalLongitude:-85.373; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:20-Mar-2003; individualID:DHJPAR0022928; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022928; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:03-SRNP-5624; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Camino Rio Francia; verbatimElevation:410; verbatimLatitude:10.904; verbatimLongitude:-85.287; verbatimCoordinateSystem:Decimal; decimalLatitude:10.904; decimalLongitude:-85.287; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:23-Dec-2004; individualID:DHJPAR0022929; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022929; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-42789; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Quebrada Lajosa; verbatimElevation:400; verbatimLatitude:11.033; verbatimLongitude:-85.429; verbatimCoordinateSystem:Decimal; decimalLatitude:11.033; decimalLongitude:-85.429; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:22-Feb-2005; individualID:DHJPAR0022930; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022930; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-20280; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Cuesta Caimito; verbatimElevation:640; verbatimLatitude:10.891; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.891; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:20-Aug-2005; individualID:DHJPAR0022931; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022931; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-46753; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Estacion Gongora; verbatimElevation:570; verbatimLatitude:10.887; verbatimLongitude:-85.474; verbatimCoordinateSystem:Decimal; decimalLatitude:10.887; decimalLongitude:-85.474; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:01-Aug-2005; individualID:DHJPAR0022932; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022932; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-46479; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Sendero Huerta; verbatimElevation:527; verbatimLatitude:10.931; verbatimLongitude:-85.372; verbatimCoordinateSystem:Decimal; decimalLatitude:10.931; decimalLongitude:-85.372; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:28-Aug-2005; individualID:DHJPAR0022933; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022933; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-4521; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Gongora Bananal; verbatimElevation:600; verbatimLatitude:10.889; verbatimLongitude:-85.476; verbatimCoordinateSystem:Decimal; decimalLatitude:10.889; decimalLongitude:-85.476; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:27-Sep-2004; individualID:DHJPAR0022934; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022934; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-48258; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Quebrada Otilio; verbatimElevation:550; verbatimLatitude:10.89; verbatimLongitude:-85.48; verbatimCoordinateSystem:Decimal; decimalLatitude:10.89; decimalLongitude:-85.48; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:31-Aug-2004; individualID:DHJPAR0022935; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022935; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-47696; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Puente Palma; verbatimElevation:460; verbatimLatitude:10.916; verbatimLongitude:-85.379; verbatimCoordinateSystem:Decimal; decimalLatitude:10.916; decimalLongitude:-85.379; samplingProtocol:reared from caterpillar of Talides Burns02 (Hesperiidae); verbatimEventDate:16-Dec-2004; individualID:DHJPAR0022936; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022936; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-60869; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Casa Roberto; verbatimElevation:520; verbatimLatitude:11.011; verbatimLongitude:-85.421; verbatimCoordinateSystem:Decimal; decimalLatitude:11.011; decimalLongitude:-85.421; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:06-Feb-2005; individualID:DHJPAR0022937; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022937; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-56649; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Sendero Pinyal; verbatimElevation:630; verbatimLatitude:10.872; verbatimLongitude:-85.393; verbatimCoordinateSystem:Decimal; decimalLatitude:10.872; decimalLongitude:-85.393; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:05-May-2005; individualID:DHJPAR0022938; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022938; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-2168; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Tangelo; verbatimElevation:410; verbatimLatitude:11.018; verbatimLongitude:-85.45; verbatimCoordinateSystem:Decimal; decimalLatitude:11.018; decimalLongitude:-85.45; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:30-Mar-2005; individualID:DHJPAR0022939; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022939; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-21176; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Quebrada Otilio; verbatimElevation:550; verbatimLatitude:10.89; verbatimLongitude:-85.48; verbatimCoordinateSystem:Decimal; decimalLatitude:10.89; decimalLongitude:-85.48; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:14-Sep-2004; individualID:DHJPAR0022940; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022940; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-48022; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Quebrada Heliconia; verbatimElevation:390; verbatimLatitude:10.886; verbatimLongitude:-85.492; verbatimCoordinateSystem:Decimal; decimalLatitude:10.886; decimalLongitude:-85.492; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:05-Oct-2004; individualID:DHJPAR0022941; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022941; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-48423; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Quebrada Otilio; verbatimElevation:550; verbatimLatitude:10.89; verbatimLongitude:-85.48; verbatimCoordinateSystem:Decimal; decimalLatitude:10.89; decimalLongitude:-85.48; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:07-Sep-2004; individualID:DHJPAR0022942; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022942; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-48023; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Quebrada Otilio; verbatimElevation:550; verbatimLatitude:10.89; verbatimLongitude:-85.48; verbatimCoordinateSystem:Decimal; decimalLatitude:10.89; decimalLongitude:-85.48; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:06-Sep-2004; individualID:DHJPAR0022943; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022943; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-48015; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Gongora Bananal; verbatimElevation:600; verbatimLatitude:10.889; verbatimLongitude:-85.476; verbatimCoordinateSystem:Decimal; decimalLatitude:10.889; decimalLongitude:-85.476; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:26-Sep-2004; individualID:DHJPAR0022944; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022944; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-48263; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Cuesta Caimito; verbatimElevation:640; verbatimLatitude:10.891; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.891; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides Burns02 (Hesperiidae); verbatimEventDate:01-Sep-2004; individualID:DHJPAR0022945; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022945; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-47944; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Estacion Pitilla; verbatimElevation:675; verbatimLatitude:10.989; verbatimLongitude:-85.426; verbatimCoordinateSystem:Decimal; decimalLatitude:10.989; decimalLongitude:-85.426; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:22-Sep-2004; individualID:DHJPAR0022946; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022946; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-34306; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Gongora Bananal; verbatimElevation:600; verbatimLatitude:10.889; verbatimLongitude:-85.476; verbatimCoordinateSystem:Decimal; decimalLatitude:10.889; decimalLongitude:-85.476; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:17-Aug-2004; individualID:DHJPAR0022947; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022947; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-47476; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Quebrada Otilio; verbatimElevation:550; verbatimLatitude:10.89; verbatimLongitude:-85.48; verbatimCoordinateSystem:Decimal; decimalLatitude:10.89; decimalLongitude:-85.48; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:10-Sep-2004; individualID:DHJPAR0022948; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022948; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-48018; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Puente Gongora; verbatimElevation:540; verbatimLatitude:10.885; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.885; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:15-Sep-2005; individualID:DHJPAR0022949; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022949; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-47227; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Vado Rio Cucaracho; verbatimElevation:640; verbatimLatitude:10.87; verbatimLongitude:-85.392; verbatimCoordinateSystem:Decimal; decimalLatitude:10.87; decimalLongitude:-85.392; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:27-Sep-2004; individualID:DHJPAR0022950; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022950; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-4450; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Quebrada Otilio; verbatimElevation:550; verbatimLatitude:10.89; verbatimLongitude:-85.48; verbatimCoordinateSystem:Decimal; decimalLatitude:10.89; decimalLongitude:-85.48; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:13-Sep-2004; individualID:DHJPAR0022951; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022951; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-48014; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Gongora Bananal; verbatimElevation:600; verbatimLatitude:10.889; verbatimLongitude:-85.476; verbatimCoordinateSystem:Decimal; decimalLatitude:10.889; decimalLongitude:-85.476; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:10-Oct-2004; individualID:DHJPAR0022952; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0022952; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-48856; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Gongora Bananal; verbatimElevation:600; verbatimLatitude:10.889; verbatimLongitude:-85.476; verbatimCoordinateSystem:Decimal; decimalLatitude:10.889; decimalLongitude:-85.476; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:16-Aug-2004; individualID:DHJPAR0023084; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0023084; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-47275; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Bosque Aguirre; verbatimElevation:620; verbatimLatitude:11.001; verbatimLongitude:-85.438; verbatimCoordinateSystem:Decimal; decimalLatitude:11.001; decimalLongitude:-85.438; samplingProtocol:reared from caterpillar of Talides Burns04 (Hesperiidae); verbatimEventDate:04-Dec-2007; individualID:DHJPAR0023086; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0023086; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-24167; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Brasilia; verbatimLocality:Piedrona; verbatimElevation:340; verbatimLatitude:11.016; verbatimLongitude:-85.359; verbatimCoordinateSystem:Decimal; decimalLatitude:11.016; decimalLongitude:-85.359; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:11-Dec-2007; individualID:DHJPAR0023665; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0023665; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-65945; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Sendero Juntas; verbatimElevation:400; verbatimLatitude:10.907; verbatimLongitude:-85.288; verbatimCoordinateSystem:Decimal; decimalLatitude:10.907; decimalLongitude:-85.288; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:03-Mar-2008; individualID:DHJPAR0027837; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0027837; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-40369; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Mundo Nuevo; verbatimLocality:Vado Huacas; verbatimElevation:490; verbatimLatitude:10.755; verbatimLongitude:-85.391; verbatimCoordinateSystem:Decimal; decimalLatitude:10.755; decimalLongitude:-85.391; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:26-Aug-2008; individualID:DHJPAR0027838; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0027838; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-57067; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Mundo Nuevo; verbatimLocality:Vado Huacas; verbatimElevation:490; verbatimLatitude:10.755; verbatimLongitude:-85.391; verbatimCoordinateSystem:Decimal; decimalLatitude:10.755; decimalLongitude:-85.391; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:27-Aug-2008; individualID:DHJPAR0029642; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0029642; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-57068; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Finca San Gabriel; verbatimElevation:645; verbatimLatitude:10.878; verbatimLongitude:-85.393; verbatimCoordinateSystem:Decimal; decimalLatitude:10.878; decimalLongitude:-85.393; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:21-Oct-2008; individualID:DHJPAR0029715; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0029715; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-5240; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Uncaria; verbatimElevation:370; verbatimLatitude:11.018; verbatimLongitude:-85.474; verbatimCoordinateSystem:Decimal; decimalLatitude:11.018; decimalLongitude:-85.474; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:09-Sep-2008; individualID:DHJPAR0029717; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0029717; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-22738; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Bosque Aguirre; verbatimElevation:620; verbatimLatitude:11.001; verbatimLongitude:-85.438; verbatimCoordinateSystem:Decimal; decimalLatitude:11.001; decimalLongitude:-85.438; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:11-Sep-2008; individualID:DHJPAR0029719; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0029719; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-22720; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Bosque Aguirre; verbatimElevation:620; verbatimLatitude:11.001; verbatimLongitude:-85.438; verbatimCoordinateSystem:Decimal; decimalLatitude:11.001; decimalLongitude:-85.438; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:12-Sep-2008; individualID:DHJPAR0029720; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0029720; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-22723; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Bosque Aguirre; verbatimElevation:620; verbatimLatitude:11.001; verbatimLongitude:-85.438; verbatimCoordinateSystem:Decimal; decimalLatitude:11.001; decimalLongitude:-85.438; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:11-Sep-2008; individualID:DHJPAR0029721; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0029721; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-22720; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Finca San Gabriel; verbatimElevation:645; verbatimLatitude:10.878; verbatimLongitude:-85.393; verbatimCoordinateSystem:Decimal; decimalLatitude:10.878; decimalLongitude:-85.393; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:28-Oct-2008; individualID:DHJPAR0029985; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0029985; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-5243; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Finca San Gabriel; verbatimElevation:645; verbatimLatitude:10.878; verbatimLongitude:-85.393; verbatimCoordinateSystem:Decimal; decimalLatitude:10.878; decimalLongitude:-85.393; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:29-Oct-2008; individualID:DHJPAR0029988; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0029988; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-5242; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Sendero Huerta; verbatimElevation:527; verbatimLatitude:10.931; verbatimLongitude:-85.372; verbatimCoordinateSystem:Decimal; decimalLatitude:10.931; decimalLongitude:-85.372; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:08-Nov-2008; individualID:DHJPAR0029994; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0029994; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-5522; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Sendero Huerta; verbatimElevation:527; verbatimLatitude:10.931; verbatimLongitude:-85.372; verbatimCoordinateSystem:Decimal; decimalLatitude:10.931; decimalLongitude:-85.372; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:07-Jan-2009; individualID:DHJPAR0030016; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0030016; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-6840; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Rio Francia Arriba; verbatimElevation:400; verbatimLatitude:10.897; verbatimLongitude:-85.29; verbatimCoordinateSystem:Decimal; decimalLatitude:10.897; decimalLongitude:-85.29; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:16-Feb-2009; individualID:DHJPAR0030034; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0030034; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-40082; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Brasilia; verbatimLocality:Gallinazo; verbatimElevation:360; verbatimLatitude:11.018; verbatimLongitude:-85.372; verbatimCoordinateSystem:Decimal; decimalLatitude:11.018; decimalLongitude:-85.372; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:15-Dec-2008; individualID:DHJPAR0030044; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0030044; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-66133; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Manguera; verbatimElevation:470; verbatimLatitude:10.996; verbatimLongitude:-85.398; verbatimCoordinateSystem:Decimal; decimalLatitude:10.996; decimalLongitude:-85.398; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:12-Nov-2008; individualID:DHJPAR0030172; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0030172; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-72761; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Bosque Aguirre; verbatimElevation:620; verbatimLatitude:11.001; verbatimLongitude:-85.438; verbatimCoordinateSystem:Decimal; decimalLatitude:11.001; decimalLongitude:-85.438; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:16-Feb-2009; individualID:DHJPAR0030192; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0030192; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-20174; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Puente Quebrada Trigal; verbatimElevation:290; verbatimLatitude:11.027; verbatimLongitude:-85.495; verbatimCoordinateSystem:Decimal; decimalLatitude:11.027; decimalLongitude:-85.495; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:30-Nov-2008; individualID:DHJPAR0030207; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0030207; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-24204; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Finca San Gabriel; verbatimElevation:645; verbatimLatitude:10.878; verbatimLongitude:-85.393; verbatimCoordinateSystem:Decimal; decimalLatitude:10.878; decimalLongitude:-85.393; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:21-Oct-2008; individualID:DHJPAR0030233; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0030233; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-5240; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Puente Palma; verbatimElevation:460; verbatimLatitude:10.916; verbatimLongitude:-85.379; verbatimCoordinateSystem:Decimal; decimalLatitude:10.916; decimalLongitude:-85.379; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:10-Mar-2009; individualID:DHJPAR0030427; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0030427; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-593; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Finca San Gabriel; verbatimElevation:645; verbatimLatitude:10.878; verbatimLongitude:-85.393; verbatimCoordinateSystem:Decimal; decimalLatitude:10.878; decimalLongitude:-85.393; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:29-Oct-2008; individualID:DHJPAR0034326; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034326; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-5242; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Sendero Huerta; verbatimElevation:527; verbatimLatitude:10.931; verbatimLongitude:-85.372; verbatimCoordinateSystem:Decimal; decimalLatitude:10.931; decimalLongitude:-85.372; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:08-Nov-2008; individualID:DHJPAR0034332; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034332; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-5522; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Finca San Gabriel; verbatimElevation:645; verbatimLatitude:10.878; verbatimLongitude:-85.393; verbatimCoordinateSystem:Decimal; decimalLatitude:10.878; decimalLongitude:-85.393; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:28-Oct-2008; individualID:DHJPAR0034342; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034342; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-5243; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Brasilia; verbatimLocality:Gallinazo; verbatimElevation:360; verbatimLatitude:11.018; verbatimLongitude:-85.372; verbatimCoordinateSystem:Decimal; decimalLatitude:11.018; decimalLongitude:-85.372; samplingProtocol:reared from caterpillar of Talides Burns (Hesperiidae); verbatimEventDate:18-Mar-2009; individualID:DHJPAR0034497; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034497; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-65324; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Rio Blanco Abajo; verbatimElevation:500; verbatimLatitude:10.9; verbatimLongitude:-85.373; verbatimCoordinateSystem:Decimal; decimalLatitude:10.9; decimalLongitude:-85.373; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:16-Jul-2006; individualID:DHJPAR0036458; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0036458; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-4769; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Corrales Viejos; verbatimElevation:495; verbatimLatitude:10.9; verbatimLongitude:-85.381; verbatimCoordinateSystem:Decimal; decimalLatitude:10.9; decimalLongitude:-85.381; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:08-Dec-2005; individualID:DHJPAR0038602; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0038602; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-7124; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Bosque Aguirre; verbatimElevation:620; verbatimLatitude:11.0006; verbatimLongitude:-85.438; verbatimCoordinateSystem:Decimal; decimalLatitude:11.0006; decimalLongitude:-85.438; samplingProtocol:reared from caterpillar of Thracides phidon (Hesperiidae); verbatimEventDate:23-Dec-2010; individualID:DHJPAR0042322; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0042322; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:10-SRNP-22751; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Jacobo; verbatimElevation:461; verbatimLatitude:10.941; verbatimLongitude:-85.318; verbatimCoordinateSystem:Decimal; decimalLatitude:10.941; decimalLongitude:-85.318; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:14-Apr-2011; individualID:DHJPAR0042704; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0042704; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:11-SRNP-69824; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Sendero Perdido; verbatimElevation:620; verbatimLatitude:10.8794; verbatimLongitude:-85.38607; verbatimCoordinateSystem:Decimal; decimalLatitude:10.8794; decimalLongitude:-85.38607; samplingProtocol:reared from caterpillar of Talides sinois (Hesperiidae); verbatimEventDate:22-Jul-2012; individualID:DHJPAR0049601; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0049601; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:12-SRNP-2735; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector San Cristobal; verbatimLocality:Sendero Huerta; verbatimElevation:527; verbatimLatitude:10.9305; verbatimLongitude:-85.37223; verbatimCoordinateSystem:Decimal; decimalLatitude:10.9305; decimalLongitude:-85.37223; samplingProtocol:reared from caterpillar of Talides Burns (Hesperiidae); verbatimEventDate:31-Oct-2011; individualID:DHJPAR0046411; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0046411; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:11-SRNP-4237; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Camino Albergue Oscar; verbatimElevation:560; verbatimLatitude:10.87741; verbatimLongitude:-85.32363; verbatimCoordinateSystem:Decimal; decimalLatitude:10.87741; decimalLongitude:-85.32363; samplingProtocol:reared from caterpillar of Talides Burns (Hesperiidae); verbatimEventDate:17-Dec-2011; individualID:DHJPAR0046465; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0046465; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:11-SRNP-5052; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Medrano; verbatimElevation:380; verbatimLatitude:11.01602; verbatimLongitude:-85.38053; verbatimCoordinateSystem:Decimal; decimalLatitude:11.01602; decimalLongitude:-85.38053; samplingProtocol:reared from caterpillar of Talides Burns02 (Hesperiidae); verbatimEventDate:07-Dec-2011; individualID:DHJPAR0046584; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0046584; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:11-SRNP-72686; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Palomo; verbatimElevation:96; verbatimLatitude:10.96187; verbatimLongitude:-85.28045; verbatimCoordinateSystem:Decimal; decimalLatitude:10.96187; decimalLongitude:-85.28045; samplingProtocol:reared from caterpillar of Talides Burns03 (Hesperiidae); verbatimEventDate:22-Nov-2011; individualID:DHJPAR0046683; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0046683; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:11-SRNP-67847; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Medrano; verbatimElevation:380; verbatimLatitude:11.01602; verbatimLongitude:-85.38053; verbatimCoordinateSystem:Decimal; decimalLatitude:11.01602; decimalLongitude:-85.38053; samplingProtocol:reared from caterpillar of Talides sergestus (Hesperiidae); verbatimEventDate:12-Feb-2012; individualID:DHJPAR0048579; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0048579; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:12-SRNP-70340; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia marioburgosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:marioburgosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Medrano; verbatimElevation:380; verbatimLatitude:11.01602; verbatimLongitude:-85.38053; verbatimCoordinateSystem:Decimal; decimalLatitude:11.01602; decimalLongitude:-85.38053; samplingProtocol:reared from caterpillar of Talides Burns03 (Hesperiidae); verbatimEventDate:20-Dec-2012; individualID:DHJPAR0050510; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0050510; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:12-SRNP-72629; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
Description
Male (Fig.






Female (Fig.
Diagnosis
Differs from S. cerussata, who also possesses fine white hairs on parafacialia, due to its broadly yellow side coloration, and yellow orange antenna.
Etymology
This species is named in honor of Sr. Mario Burgos Cespedes (RIP), whose large land holdings in Sector Pocosol became major parts of the newly forming Area de Conservacion Guanacaste in the late 1980's.
Distribution
Costa Rica, ACG, Prov. Alajuela and Guanacaste, rain forest and dry forest, 96–1140 m elevation.
Ecology
Reared 221 times from 5 species of Talides (Hesperiidae; Hesperiinae), all feeding on broad-leafed rain forest monocots (Heliconia spp., Heliconiaceae and Musa spp., introduced Musaceae). Usually 5–12 larvae per caterpillar. This frequently-reared species of Spathidexia displays a very shallow DNA barcode split into two groups (see Supplemental Appendix 1) that are identical in morphology, microgeographic location in ACG, and host records. We judge these specimens to be one species of Spathidexia but they may merit a subsequent deeper genetic probe.
Spathidexia luisrobertogallegosi , sp. n.
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:15-Apr-2009; individualID:DHJPAR0034394; individualCount:1; sex:M; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034394; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35179; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Puente Gongora; verbatimElevation:540; verbatimLatitude:10.885; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.885; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Vehilius illudens (Hesperiidae); verbatimEventDate:22-Nov-2006; individualID:DHJPAR0016675; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0016675; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-47851; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Puente Gongora; verbatimElevation:540; verbatimLatitude:10.885; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.885; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:19-Feb-2007; individualID:DHJPAR0017097; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0017097; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-45135; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Quebrada Trigal; verbatimElevation:290; verbatimLatitude:11.027; verbatimLongitude:-85.495; verbatimCoordinateSystem:Decimal; decimalLatitude:11.027; decimalLongitude:-85.495; samplingProtocol:reared from caterpillar of Nyctelius nyctelius (Hesperiidae); verbatimEventDate:07-Feb-2005; individualID:DHJPAR0018737; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0018737; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:05-SRNP-20039; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Quebrada Raiz; verbatimElevation:280; verbatimLatitude:11.029; verbatimLongitude:-85.487; verbatimCoordinateSystem:Decimal; decimalLatitude:11.029; decimalLongitude:-85.487; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:07-Jun-2004; individualID:DHJPAR0018738; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0018738; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-22017; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Del Oro; verbatimLocality:Canyon Rio Mena; verbatimElevation:560; verbatimLatitude:10.996; verbatimLongitude:-85.456; verbatimCoordinateSystem:Decimal; decimalLatitude:10.996; decimalLongitude:-85.456; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:08-Mar-2002; individualID:DHJPAR0018740; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0018740; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:02-SRNP-5601; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:20-Apr-2007; individualID:DHJPAR0019615; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019615; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-35371; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Arenales; verbatimElevation:1080; verbatimLatitude:10.925; verbatimLongitude:-85.467; verbatimCoordinateSystem:Decimal; decimalLatitude:10.925; decimalLongitude:-85.467; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:12-Apr-2007; individualID:DHJPAR0019623; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019623; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-35277; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:12-Apr-2007; individualID:DHJPAR0019633; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019633; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-35362; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Estacion Cacao; verbatimElevation:1150; verbatimLatitude:10.927; verbatimLongitude:-85.468; verbatimCoordinateSystem:Decimal; decimalLatitude:10.927; decimalLongitude:-85.468; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:14-May-2007; individualID:DHJPAR0019648; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019648; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-35646; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:05-May-2007; individualID:DHJPAR0019649; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019649; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-35454; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:01-May-2007; individualID:DHJPAR0019650; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019650; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-35357; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:03-Apr-2007; individualID:DHJPAR0019651; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019651; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-35243; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Cerro Pedregal; verbatimElevation:1080; verbatimLatitude:10.928; verbatimLongitude:-85.474; verbatimCoordinateSystem:Decimal; decimalLatitude:10.928; decimalLongitude:-85.474; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:14-Apr-2007; individualID:DHJPAR0019652; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019652; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-35297; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:18-Apr-2007; individualID:DHJPAR0019653; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019653; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-35367; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Cima; verbatimElevation:1460; verbatimLatitude:10.933; verbatimLongitude:-85.457; verbatimCoordinateSystem:Decimal; decimalLatitude:10.933; decimalLongitude:-85.457; samplingProtocol:reared from caterpillar of hespJanzen01 Janzen38 (Hesperiidae); verbatimEventDate:30-May-2007; individualID:DHJPAR0019654; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019654; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-35729; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:09-Apr-2007; individualID:DHJPAR0019655; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019655; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-35375; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Arenales; verbatimElevation:1080; verbatimLatitude:10.925; verbatimLongitude:-85.467; verbatimCoordinateSystem:Decimal; decimalLatitude:10.925; decimalLongitude:-85.467; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:14-Apr-2007; individualID:DHJPAR0019656; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019656; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-35335; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:20-Apr-2007; individualID:DHJPAR0019657; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019657; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-35369; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Cerro Pedregal; verbatimElevation:1080; verbatimLatitude:10.928; verbatimLongitude:-85.474; verbatimCoordinateSystem:Decimal; decimalLatitude:10.928; decimalLongitude:-85.474; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:18-Apr-2007; individualID:DHJPAR0019659; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019659; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-35298; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:23-Apr-2007; individualID:DHJPAR0019660; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0019660; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:07-SRNP-35372; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:05-May-2009; individualID:DHJPAR0034378; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034378; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35196; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:18-Apr-2009; individualID:DHJPAR0034380; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034380; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35197; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:20-Apr-2009; individualID:DHJPAR0034381; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034381; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35184; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Arenales; verbatimElevation:1080; verbatimLatitude:10.925; verbatimLongitude:-85.467; verbatimCoordinateSystem:Decimal; decimalLatitude:10.925; decimalLongitude:-85.467; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:27-Sep-2009; individualID:DHJPAR0034382; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034382; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35251; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:15-Apr-2009; individualID:DHJPAR0034383; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034383; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35178; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:24-Apr-2009; individualID:DHJPAR0034384; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034384; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35182; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:20-Apr-2009; individualID:DHJPAR0034385; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034385; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35190; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:27-Sep-2009; individualID:DHJPAR0034386; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034386; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35183; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:27-Sep-2009; individualID:DHJPAR0034388; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034388; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35198; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:18-Apr-2009; individualID:DHJPAR0034389; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034389; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35187; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:23-Apr-2009; individualID:DHJPAR0034390; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034390; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35194; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:27-Sep-2009; individualID:DHJPAR0034391; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034391; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35189; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Arenales; verbatimElevation:1080; verbatimLatitude:10.925; verbatimLongitude:-85.467; verbatimCoordinateSystem:Decimal; decimalLatitude:10.925; decimalLongitude:-85.467; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:27-Sep-2009; individualID:DHJPAR0034392; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034392; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35202; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.924; verbatimLongitude:-85.47; verbatimCoordinateSystem:Decimal; decimalLatitude:10.924; decimalLongitude:-85.47; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:12-Apr-2009; individualID:DHJPAR0034393; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034393; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35192; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Sendero Nacho; verbatimElevation:710; verbatimLatitude:10.984; verbatimLongitude:-85.425; verbatimCoordinateSystem:Decimal; decimalLatitude:10.984; decimalLongitude:-85.425; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:13-Apr-2011; individualID:DHJPAR0042574; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0042574; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:11-SRNP-30498; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Sendero Laguna; verbatimElevation:680; verbatimLatitude:10.989; verbatimLongitude:-85.423; verbatimCoordinateSystem:Decimal; decimalLatitude:10.989; decimalLongitude:-85.423; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:18-Apr-2011; individualID:DHJPAR0042576; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0042576; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:11-SRNP-30640; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Sendero Laguna; verbatimElevation:680; verbatimLatitude:10.989; verbatimLongitude:-85.423; verbatimCoordinateSystem:Decimal; decimalLatitude:10.989; decimalLongitude:-85.423; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:07-Dec-2011; individualID:DHJPAR0048559; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0048559; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:11-SRNP-33677; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Loaiciga; verbatimElevation:445; verbatimLatitude:11.01983; verbatimLongitude:-85.41342; verbatimCoordinateSystem:Decimal; decimalLatitude:11.01983; decimalLongitude:-85.41342; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:04-Jan-2012; individualID:DHJPAR0048614; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0048614; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:12-SRNP-30000; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Ingas; verbatimElevation:580; verbatimLatitude:11.00311; verbatimLongitude:-85.42041; verbatimCoordinateSystem:Decimal; decimalLatitude:11.00311; decimalLongitude:-85.42041; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:07-Jan-2012; individualID:DHJPAR0048564; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0048564; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:12-SRNP-30066; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Pasmompa; verbatimElevation:440; verbatimLatitude:11.01926; verbatimLongitude:-85.40997; verbatimCoordinateSystem:Decimal; decimalLatitude:11.01926; decimalLongitude:-85.40997; samplingProtocol:reared from caterpillar of Justinia norda (Hesperiidae); verbatimEventDate:04-Jan-2012; individualID:DHJPAR0048608; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0048608; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:12-SRNP-30209; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Sendero Rotulo; verbatimElevation:510; verbatimLatitude:11.01355; verbatimLongitude:-85.42406; verbatimCoordinateSystem:Decimal; decimalLatitude:11.01355; decimalLongitude:-85.42406; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:31-Jan-2012; individualID:DHJPAR0048609; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0048609; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:12-SRNP-30302; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Pasmompa; verbatimElevation:440; verbatimLatitude:11.01926; verbatimLongitude:-85.40997; verbatimCoordinateSystem:Decimal; decimalLatitude:11.01926; decimalLongitude:-85.40997; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:26-Feb-2012; individualID:DHJPAR0048629; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0048629; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:12-SRNP-30522; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Ingas; verbatimElevation:580; verbatimLatitude:11.00311; verbatimLongitude:-85.42041; verbatimCoordinateSystem:Decimal; decimalLatitude:11.00311; decimalLongitude:-85.42041; samplingProtocol:reared from caterpillar of Anthoptus epictetus (Hesperiidae); verbatimEventDate:15-Feb-2012; individualID:DHJPAR0048603; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0048603; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:12-SRNP-30526; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Estacion Pitilla; verbatimElevation:675; verbatimLatitude:10.98931; verbatimLongitude:-85.42581; verbatimCoordinateSystem:Decimal; decimalLatitude:10.98931; decimalLongitude:-85.42581; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:27-Mar-2012; individualID:DHJPAR0048636; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0048636; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:12-SRNP-30636; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luisrobertogallegosi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luisrobertogallegosi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Sendero Nayo; verbatimElevation:1090; verbatimLatitude:10.92446; verbatimLongitude:-85.46953; verbatimCoordinateSystem:Decimal; decimalLatitude:10.92446; decimalLongitude:-85.46953; samplingProtocol:reared from caterpillar of Conga chydea (Hesperiidae); verbatimEventDate:18-Feb-2009; individualID:DHJPAR0034394; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0034394; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:09-SRNP-35179; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
Description
Male (Fig.






Female (Fig.
Diagnosis
Differs from S. cerussata, who also possesses fine white hairs on parafacialia, due to its gold thorax, broadly yellow lateral coloration, and yellow orange antenna with darkened apices.
Etymology
This species is named in honor of Sr. Luis Roberto Gallegos (RIP), whose large land holdings in Sector El Hacha, Sector Orosi and Sector Pocosol became major parts of the newly forming Area de Conservacion Guanacaste in the late 1980's.
Distribution
Costa Rica, ACG, Prov. Guanacaste, rain forest, cloud forest (marginally) and dry forest-rain forest interface (primarily), 280–1460 m elevation.
Ecology
Reared 50 times from Conga chydea (Hesperiidae, Hesperiinae) only, always feeding on grasses on forest edges in rain forest and the interface between rain forest and dry forest.
Spathidexia luteola , sp. n.
-
scientificName: Spathidexia luteola; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luteola; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Gongora Bananal; verbatimElevation:600; verbatimLatitude:10.88919; verbatimLongitude:-85.47609; verbatimCoordinateSystem:Decimal; decimalLatitude:10.88919; decimalLongitude:-85.47609; samplingProtocol:reared from caterpillar of Vacerra aeas (Hesperiidae); verbatimEventDate:03-Apr-2004; individualID:04-SRNP-45008; individualCount:1; sex:M; lifeStage:adult; preparations:pinned; catalogNumber:04-SRNP-45008; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luteola; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luteola; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Santa Rosa; verbatimLocality:Bosque San Emilio; verbatimElevation:300; verbatimLatitude:10.84389; verbatimLongitude:-85.61384; verbatimCoordinateSystem:Decimal; decimalLatitude:10.84389; decimalLongitude:-85.61384; samplingProtocol:reared from caterpillar of Moeris stroma (Hesperiidae); verbatimEventDate:22-Oct-2004; individualID:DHJPAR0011066; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0011066; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-14634; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luteola; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luteola; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Gongora Bananal; verbatimElevation:600; verbatimLatitude:10.88919; verbatimLongitude:-85.47609; verbatimCoordinateSystem:Decimal; decimalLatitude:10.88919; decimalLongitude:-85.47609; samplingProtocol:reared from caterpillar of Vacerra aeas (Hesperiidae); verbatimEventDate:03-Apr-2004; individualID:DHJPAR0018741; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0018741; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-45008; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luteola; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luteola; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Gongora Bananal; verbatimElevation:600; verbatimLatitude:10.88919; verbatimLongitude:-85.47609; verbatimCoordinateSystem:Decimal; decimalLatitude:10.88919; decimalLongitude:-85.47609; samplingProtocol:reared from caterpillar of Vacerra aeas (Hesperiidae); verbatimEventDate:03-Apr-2004; individualID:DHJPAR0018742; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0018742; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-45008; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia luteola; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:luteola; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Gongora Bananal; verbatimElevation:600; verbatimLatitude:10.88919; verbatimLongitude:-85.47609; verbatimCoordinateSystem:Decimal; decimalLatitude:10.88919; decimalLongitude:-85.47609; samplingProtocol:reared from caterpillar of Vacerra aeas (Hesperiidae); verbatimEventDate:03-Apr-2004; individualID:04-SRNP-45008; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:04-SRNP-45008; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
Description
Male (Fig.

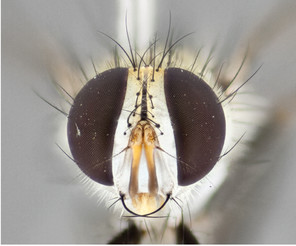




Female (Fig.
Diagnosis
Species closely resembling S. clemonsi, differs from the former due to the presence of yellow tibiae on the front pair of legs, yellow/pale hairs covering the entire body, and costal spine is reduced in S. luteola, not absent as in S. clemonsi. This species is quite similar to Spathidexia brasiliensis, from which it can be differentiated by the former having an orange pedicel and narrow ocellar triangle.
Etymology
From the Latin adjective, “luteus”, yellow, in reference to the pale yellow hairs covering the body of this species, particularly prominent on the underside of the abdomen.
Distribution
Costa Rica, ACG, Prov. Guanacaste, dry forest and dry forest-rain forest interface, 300–600 m elevation.
Ecology
Reared from 2 species of Hesperiidae, Hesperiinae, eating Lasiacis (Poaceae) (2 records), bearing 1–5 larvae per caterpillar.
Spathidexia hernanrodriguezi , sp. n.
-
scientificName: Spathidexia hernanrodriguezi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:hernanrodriguezi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pocosol; verbatimLocality:Quebrada Aserradero; verbatimElevation:160; verbatimLatitude:10.899; verbatimLongitude:-85.564; verbatimCoordinateSystem:Decimal; decimalLatitude:10.899; decimalLongitude:-85.564; samplingProtocol:reared from caterpillar of Corticea corticea (Hesperiidae); verbatimEventDate:24-Apr-2006; individualID:DHJPAR0018743; individualCount:1; sex:M; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0018743; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-10616; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia hernanrodriguezi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:hernanrodriguezi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Puente Gongora; verbatimElevation:540; verbatimLatitude:10.885; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.885; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Quasimellana antipazina (Hesperiidae); verbatimEventDate:02-May-2004; individualID:DHJPAR0010022; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0010022; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-45170; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia hernanrodriguezi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:hernanrodriguezi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pocosol; verbatimLocality:Quebrada Aserradero; verbatimElevation:160; verbatimLatitude:10.899; verbatimLongitude:-85.564; verbatimCoordinateSystem:Decimal; decimalLatitude:10.899; decimalLongitude:-85.564; samplingProtocol:reared from caterpillar of Corticea corticea (Hesperiidae); verbatimEventDate:02-May-2004; individualID:DHJPAR0018744; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0018744; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:04-SRNP-10616; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia hernanrodriguezi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:hernanrodriguezi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Cacao; verbatimLocality:Puente Gongora; verbatimElevation:540; verbatimLatitude:10.885; verbatimLongitude:-85.472; verbatimCoordinateSystem:Decimal; decimalLatitude:10.885; decimalLongitude:-85.472; samplingProtocol:reared from caterpillar of Quasimellana antipazina (Hesperiidae); verbatimEventDate:24-Apr-2006; individualID:DHJPAR0018745; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0018745; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-45170; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia hernanrodriguezi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:hernanrodriguezi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pocosol; verbatimLocality:Pozo Centeno; verbatimElevation:155; verbatimLatitude:10.9; verbatimLongitude:-85.566; verbatimCoordinateSystem:Decimal; decimalLatitude:10.9; decimalLongitude:-85.566; samplingProtocol:reared from caterpillar of Cymaenes odilia trebius (Hesperiidae); verbatimEventDate:22-Apr-2008; individualID:DHJPAR0024529; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0024529; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-12254; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia hernanrodriguezi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:hernanrodriguezi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pocosol; verbatimLocality:Pozo Centeno; verbatimElevation:155; verbatimLatitude:10.9; verbatimLongitude:-85.566; verbatimCoordinateSystem:Decimal; decimalLatitude:10.9; decimalLongitude:-85.566; samplingProtocol:reared from caterpillar of Vacerra aeas; verbatimEventDate:29-Apr-2008; individualID:DHJPAR0024530; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0024530; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:08-SRNP-12251; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
Description
Male (Fig.






Female (Fig.
Diagnosis
Resembles S. cerussata, differs from the former in the presence of apically enlarged palpus, thorax with almost indistinguisheable vittae, a haired wing vein R1, and orange pedicel. Females in S. cerussata are similarly colored to males a characteristic not shared by S. hernanrodriguezi.
Etymology
This species is named in honor of Sr. Hernan Rodriguez Arce (RIP), whose Finca San Gerardo became a large part of Sector San Cristobal of the newly forming Area de Conservación Guanacaste in the early 1990's.
Distribution
Costa Rica, ACG, Guanacaste, dry forest, 155–540 m elevation.
Ecology
Reared from 3 species of Hesperiidae, Hesperiinae (4 records), feeding on grasses. 5–12 larvae per caterpillar.
Spathidexia aurantiaca , sp. n.
-
scientificName: Spathidexia aurantiaca; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:aurantiaca; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Caverna; verbatimElevation:420; verbatimLatitude:10.927; verbatimLongitude:-85.288; verbatimCoordinateSystem:Decimal; decimalLatitude:10.927; decimalLongitude:-85.288; samplingProtocol:reared from caterpillars of Justinia Burns01 (Hesperiidae); verbatimEventDate:01-Aug-2006; individualID:DHJPAR0010326; individualCount:1; sex:M; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0010326; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-42439; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia aurantiaca; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:aurantiaca; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Puente Rio Negro; verbatimElevation:340; verbatimLatitude:10.904; verbatimLongitude:-85.303; verbatimCoordinateSystem:Decimal; decimalLatitude:10.904; decimalLongitude:-85.303; samplingProtocol:reared from caterpillars of Justinia Burns01 (Hesperiidae); verbatimEventDate:12-Mar-2006; individualID:DHJPAR0006934; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0006934; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-40499; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia aurantiaca; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:aurantiaca; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Puente Rio Negro; verbatimElevation:340; verbatimLatitude:10.904; verbatimLongitude:-85.303; verbatimCoordinateSystem:Decimal; decimalLatitude:10.904; decimalLongitude:-85.303; samplingProtocol:reared from caterpillars of Justinia Burns01 (Hesperiidae); verbatimEventDate:12-Mar-2006; individualID:DHJPAR0018746; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0018746; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:06-SRNP-40499; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia aurantiaca; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:aurantiaca; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Selva; verbatimElevation:410; verbatimLatitude:10.923; verbatimLongitude:-85.319; verbatimCoordinateSystem:Decimal; decimalLatitude:10.923; decimalLongitude:-85.319; samplingProtocol:reared from caterpillars of Euphyes Janzen01 (Hesperiidae); verbatimEventDate:05-Dec-2010; individualID:DHJPAR0040837; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0040837; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:10-SRNP-81062; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia aurantiaca; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:aurantiaca; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Selva; verbatimElevation:410; verbatimLatitude:10.923; verbatimLongitude:-85.319; verbatimCoordinateSystem:Decimal; decimalLatitude:10.923; decimalLongitude:-85.319; samplingProtocol:reared from caterpillars of Euphyes Janzen01 (Hesperiidae); verbatimEventDate:04-Dec-2010; individualID:DHJPAR0040838; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0040838; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:10-SRNP-81061; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia aurantiaca; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:aurantiaca; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Alajuela; county:Area de Conservacion Guanacaste; locality:Sector Rincon Rain Forest; verbatimLocality:Selva; verbatimElevation:410; verbatimLatitude:10.923; verbatimLongitude:-85.319; verbatimCoordinateSystem:Decimal; decimalLatitude:10.923; decimalLongitude:-85.319; samplingProtocol:reared from caterpillars of Euphyes Janzen01 (Hesperiidae); verbatimEventDate:06-May-2011; individualID:DHJPAR0042708; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0042708; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:11-SRNP-80003; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
Description
Male (Fig.






Female (Fig.
Diagnosis
This species differs from the only other reddish-orange species in Spathidexia, S. creolensis from the southern US in that Spathidexia aurantiaca has more pronounced pruinosity along abdominal margins, black ground color on T5 in both sexes, and gold along the fronto-orbital plate.
Etymology
From Latin, "aurantiaco" referring to the orange ground color of the abdomen.
Distribution
Costa Rica, ACG, Prov. Alajuela, rain forest, 340–420 m elevation.
Ecology
Reared from Hesperiidae, specifically two species of sedge (Cyperaceae)-eating Hesperiinae (4 records). 1–5 larvae per caterpillar.
Spathidexia juanvialesi , sp. n.
-
scientificName: Spathidexia juanvialesi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:juanvialesi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Bullas; verbatimElevation:440; verbatimLatitude:10.9867; verbatimLongitude:-85.38503; verbatimCoordinateSystem:Decimal; decimalLatitude:10.9867; decimalLongitude:-85.38503; samplingProtocol:reared from caterpillar of Cissia 95-SRNP-7306 (Nymphalidae); verbatimEventDate:6/Jan/12; individualID:DHJPAR0046610; individualCount:1; sex:F; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0046610; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:11-SRNP-72577; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
-
scientificName: Spathidexia juanvialesi; phylum:Arthropoda; class:Insecta; order:Diptera; family:Tachinidae; genus:Spathidexia; specificEpithet:juanvialesi; scientificNameAuthorship:Fleming & Wood, 2015; continent:Central America; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; county:Area de Conservacion Guanacaste; locality:Sector Pitilla; verbatimLocality:Ingas; verbatimElevation:580; verbatimLatitude:11.00311; verbatimLongitude:-85.42041; verbatimCoordinateSystem:Decimal; decimalLatitude:11.00311; decimalLongitude:-85.42041; samplingProtocol:reared from caterpillar of Hermeuptychia hermesDHJ04 (Nymphalidae); verbatimEventDate:2/Dec/11; individualID:DHJPAR0046623; individualCount:1; lifeStage:adult; preparations:pinned; catalogNumber:DHJPAR0046623; occurrenceDetails: ; recordedBy:D.H. Janzen & W. Hallwachs; otherCatalogNumbers:11-SRNP-33171; identifiedBy:AJ Fleming; dateIdentified:2014; language:en; institutionCode:CNC; collectionCode:Insects; basisOfRecord:Pinned Specimen
Description
Female(Fig.



Diagnosis
Similar to S. setipennis in the thoracic markings, however S. juanvialesi is strongly different in the coloration of the antenna, and the extent of the yellow on the legs.
Etymology
This species is named in honor of Sr. Juan Viales (RIP), whose Finca San Cristobal became a large part of Sector San Cristobal of the newly forming Area de Conservación Guanacaste in the eary 1990's.
Distribution
Costa Rica, ACG, Prov. Guanacaste, rain forest, 440–580 m elevation.
Ecology
Reared twice from grass-eating rain forest Hesperiinae (Hesperiidae).
Identification keys
|
Key to the species of Spathidexia reared from Area de Conservación Guanacaste, Northwestern Costa Rica This key was prepared based on the specimens collected as a result of the 40+ year rearing effort conducted in ACG. Our key is intended to speak to the fauna present within the confines of the ACG and is not meant to be an exhaustive key to the species within the confines of Costa Rica. |
||
| 1 | Proclinate orbital bristles absent | 2 |
| – | Proclinate orbital bristles present | 4 |
| 2 | Fronto-orbital plate entirely silver (no trace of gold)(Figs |
3 |
| – | Fronto-orbital plate bearing at least traces of gold at least at apex, and surrounding ocellar triangle (Figs |
Spathidexia luteola sp. n. (♂) |
| 3 | Thoracic vittae black, prominent; vein R1 bare (Fig. |
Spathidexia atripalpus sp. n. (♂) |
| – | Grey thorax with greyish indistinct vittae (only slightly visible, Fig. |
Spathidexia marioburgosi sp. n. (♂) |
| 4 | Ground color of abdomen dark, but bearing lateral orange/yellow blotches; or mostly orange (Figs |
5 |
| – | Abdominal tergites black, with grayish tomentose bands covering up to basal 1/3 of tergal surface, and with no traces of orange/yellow blotching on abdomen (Figs |
9 |
| 5 | Ground color of abdomen dark, but bearing lateral orange/yellow blotches (Figs |
6 |
| – | Ground color of abdomen orange along its entirety; or with darkened T5 in the case of females (Fig. |
Spathidexia aurantiaca sp. n. (♂ & ♀) |
| 6 | Abdomen bearing orange/yellow blotches along sides of abdominal tergites, (T1+2, T3, T4, and sometimes T5) (Figs |
7 |
| – | Abdomen with yellow blotches only on T1+2 (Fig. |
Spathidexia atripalpus sp. n. (♀) |
| 7 | Fronto-orbital plate entirely silver with no trace of gold, or if gold present, then only in trace amount alongside ocellar triangle (Figs |
8 |
| – | Fronto-orbital plate bearing traces of gold at least at apex, and surrounding ocellar triangle up to insertion of first proclinate orbital bristle (Fig. |
Spathidexia luteola sp. n. (♀) |
| 8 | Fronto-orbital plate entirely silver with no trace of gold; wide light brown frontal vitta; antenna black only orange on pedicel (Fig. |
Spathidexia juanvialesi sp. n. (♀) |
| – | Fronto-orbital plate entirely silver with slight trace of gold alongside ocellar triangle; narrow black frontal vitta; antenna bright orange (Fig. |
Spathidexia marioburgosi sp. n. (♀) |
| 9 | Fronto-orbital plate with gold color only at apex surrounding ocellar triangle, descending to insertion of upper proclinate orbital in the case of females; antenna dark, only pedicel appears orange (Fig. |
Spathidexia hernanrodriguezi sp. n. (♂ & ♀) |
| – | Fronto-orbital plate with gold color extending up to second proclinate orbital bristle; antenna orange (Fig. |
Spathidexia luisgallegosi sp. n. (♂ & ♀) |
Acknowledgements
We gratefully acknowledge the unflagging support of the team of ACG parataxonomists (Janzen et al. 2009, Janzen & Hallwachs 2011) who found and reared the specimens used in this study, and the team of biodiversity managers who protect and manage the ACG forests that host these tachinids and their caterpillar hosts. The study has been supported by U.S. National Science Foundation grants BSR 9024770 and DEB 9306296, 9400829, 9705072, 0072730, 0515699, and grants from the Wege Foundation, International Conservation Fund of Canada, Jessie B. Cox Charitable Trust, Blue Moon Fund, Guanacaste Dry Forest Conservation Fund, Area de Conservación Guanacaste, Permian Global, and University of Pennsylvania (DHJ & WH). This study has been supported by the Government of Canada through its ongoing support of the Canadian National Collection, Genome Canada, the Biodiversity Institute of Ontario, and the Ontario Genomics Institute (2008-0GI-ICI-03) (MAS), and by a Discovery Grant from Natural Sciences and Engineering Research Council of Canada (MAS). The authors also wish to acknowledge the efforts and time of the editor and reviewers, who provided insights and suggestions allowing us to make our work better.
References
- A review of the genus Spathidexia Townsend (Diptera: Tachinidae).The Wasmann Journal of Biology18:1‑36.
- A Host-Parasite Catalog of North American Tachinidae (Diptera).United States Department of Agriculture,Washington, D.C.,860pp.
- Die Zweiflügler des Kaiserlichen Museums zu Wien. V. Vorarbeiten zu einer Monographie der Muscaria Schizometopa (exclusive Anthomyidae). Pars II.Denkschriften der Kaiserlichen Akademie der Wissenschaften Wien. Mathematisch-Naturwissenschaftliche Classe58:305‑446.
- Notes and descriptions of Tachinidae.Journal of the New York Entomological Society3:49‑58.
- Revision of the Tachinidae of America north of Mexico. A family of parasitic two-winged insects.Government Printing Office,156pp. https://doi.org/10.5962/bhl.title.9264
- Annotated keys to the genera of Tachinidae (Diptera) found in tropical and southern Africa.Annals of the Natal Museum26:189‑337.
- Cumming JM, Wood DM (2009) Adult morphology and terminology. Pp. 9–50. In: Brown BV, Borkent A, Cumming JM, Wood DM, Woodley NE, Zumbado MA (Eds) Manual of Central American Diptera.1.NRC Research Press,Ottawa, Canada,xi + 714 pp.pp.
- Review of Apanteles sensu stricto (Hymenoptera, Braconidae, Microgastrinae) from Area de Conservación Guanacaste, northwestern Costa Rica, with keys to all described species from Mesoamerica.Zookeys383:1‑565. https://doi.org/10.3897/zookeys.383.6418
- Revision of the New World species of Houghia Coquillett (Diptera, Tachinidae) reared from caterpillars in Area de Conservación Guanacaste, Costa Rica.Zootaxa3858(1):1‑90. https://doi.org/10.11646/zootaxa.3858.1.1
- A new species of Cordyligaster Macquart, reared from caterpillars in Area de Conservacion Guanacaste, northwestern Costa Rica.Biodiversity Data Journal2:1‑33. URL: http://zoobank.org/A5160572-0FC6-42F1-9240-A2F4960A1E76
- Diagnosi di nuovi generi e di nuove specie di Ditteri.Bolletino dei Musei di Zoologia ed Anatomia compara della R. Università di Torino8:1‑11.
- Guimarães JH (1971) 104. Family Tachinidae. In: Papavero N (Ed.) A catalog of the Diptera of the Americas South of the United States.Museu de Zoologia, Universidade de São Paulo,São Paulo,333 p.pp.
- Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator.Proceedings of the National Academy of Sciences101(41):14812‑14817. https://doi.org/10.1073/pnas.0406166101
- An inexpensive, automation-friendly protocol for recovering high-quality DNA.Molecular Ecology Notes6(4):998‑1002. https://doi.org/10.1111/j.1471-8286.2006.01428.x
- Joining inventory by parataxonomists with DNA barcoding of a large complex tropical conserved wildland in northwestern Costa Rica.PLoS ONE6(8):1‑13. https://doi.org/10.1371/journal.pone.0018123
- Reading the complex Skipper butterfly fauna of one tropical place.PLoS ONE6(8):e19874. https://doi.org/10.1371/journal.pone.0019874
- Integration of DNA barcoding into an ongoing inventory of complex tropical biodiversity.Molecular Ecology Resources9 Suppl s1:1‑26. https://doi.org/10.1111/j.1755-0998.2009.02628.x
- Essais sur les Tachinaires (Larvaevoridae). In: Monographies publiées par les stations et laboratoires de recherches agronomiques.Ministeres de L'Agriculture, Imprimerie Nationale,Paris,1-67pp. [InFrench].
- Tachinidae (Diptera): Nomenclatural review and changes, primarily for America north of Mexico.The Canadian Entomologist130(6):751‑774. https://doi.org/10.4039/ent130751-6
- Catalogue of the Tachinidae (Diptera) of America north of Mexico.Memoirs on Entomology, International18:iv‑410.
- BOLD: The Barcode of Life Data System (http://www.barcodinglife.org).Molecular Ecology Notes7:355‑364. https://doi.org/10.1111/j.1471-8286.2007.01678.x
- North American parasitic flies of the genus Spathidexia with descriptions of two new species.Bulletin of the Brooklyn Entomological Society29:150‑154.
- New species of Tachinidae from Mexico (Diptera).Bulletin of the Brooklyn Entomological Society48:89‑96.
- New Nearctic Sarcophagidae and Tachinidae (Diptera).Bulletin of the Brooklyn Entomological Society50:122‑133.
- North American Tachinidae (Diptera).Journal of the Kansas Entomological Society37:35‑51.
- Extrapolations from field studies and known faunas converge on dramatically increased estimates of global microgastrine parasitoid wasp species richness (Hymenoptera: Braconidae).Insect Conservation and Diversity6(4):530‑536. https://doi.org/10.1111/icad.12003
- The neighbor-joining method: a new method for reconstructing phylogenetic trees.Molecular Biology and Evolution4:406‑425.
- Hyperparasitoid wasps (Hymenoptera, Trigonalidae) reared from dry forest and rain forest caterpillars of Area de Conservación Guanacaste, Costa Rica.29:119‑144. https://doi.org/10.3897/jhr.29.3233
- DNA barcodes affirm that 16 species of apparently generalist tropical parasitoid flies (Diptera: Tachinidae) are not all generalists.Proceedings of the National Academy of Sciences104:4967‑4972. https://doi.org/10.1073/pnas.0700050104
- DNA barcodes reveal cryptic host-specificity within the presumed polyphagous members of a genus of parasitoid flies (Diptera: Tachinidae).Proceedings of the National Academy of Sciences103:3657‑3662. https://doi.org/10.1073/pnas.0511318103
- Extreme diversity of tropical parasitoid wasps exposed by iterative integration of natural history, DNA barcoding, morphology, and collections.Proceedings of the National Academy of Sciences105:12359‑12364. https://doi.org/10.1073/pnas.0805319105
- The tachinids of Trinidad. II. Echinomyiines, Dexiines and allies (Dipt. Tachinidae).Canadian Journal of Zoology41:335. https://doi.org/10.1139/z63-029
- Foundation of some new genera and species of Muscoid flies mainly on reproductive and early-stage characters.Journal New York Entomological Society20:107‑119.
- Some muscoid synonyms.Entomological News26:366.
- New genera and species of muscoid flies.Proceedings of The United States National Museum56:541‑592. https://doi.org/10.5479/si.00963801.2301.541
- New muscoid genera, species and synonymy.Insecutor Inscitiae Menestruus6:157‑182.
- Synopse dos generos muscoideos da região humida tropical da America, com generos e espécies novos.Revista do Museo Paulista15:203‑385.
- New muscoid genera and species of the coasts of Perú and Chile.Revista Chilena de Historia Natural31:158‑164.
- van der Wulp FM (1890) Fam. Muscidae. Pp. 201–208. [Cont.]. In: Godman FD, Salvin O (Eds) Biologia Centrali-Americana, or, contributions to the knowledge of the fauna and flora of Mexico and Central America. Zoologia. Class Insecta. Order Diptera. Vol. 2. [1888–1903.].Taylor & Francis,London,489 pp. + 13 pls.pp.
- van der Wulp FM (1891) Fam. Muscidae. [Cont.]Pp. 209–224, 225–248, 249–264 + pls. 5-6 [cont.]. In: Godman FD, Salvin O (Eds) Biologia Centrali-Americana, or, contributions to the knowledge of the fauna and flora of Mexico and Central America. Zoologia. Class Insecta. Order Diptera. Vol. 2. [1888–1903.].Taylor & Francis,London,489 pp. + 13 pls.pp.
- Tachinidae In: Manual of Nearctic Diptera, vol. 2.Agriculture Canada Research Branch,Ottawa,77pp.
- Wood DM, Zumbado MA (2010) Tachinidae (tachinid flies, parasitic flies). Pp. 1343–1417. In: Brown BV, Borkent A, Cumming JM, Wood DM, Woodley NE, Zumbado MA (Eds) Manual of Central American Diptera. Vol 2.NRC Research Press,Ottawa, Canada,xvi + 715–1442 pp.pp.