|
Biodiversity Data Journal :
Data paper
|
|
Corresponding author:
Academic editor: Christos Arvanitidis
Received: 26 Jul 2015 | Accepted: 07 Sep 2015 | Published: 11 Sep 2015
© 2015 Emilia Rota, Yde de Jong
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Rota E, de Jong Y (2015) Fauna Europaea: Annelida - Terrestrial Oligochaeta (Enchytraeidae and Megadrili), Aphanoneura and Polychaeta. Biodiversity Data Journal 3: e5737. https://doi.org/10.3897/BDJ.3.e5737
|
|
Abstract
Fauna Europaea provides a public web-service with an index of scientific names (including important synonyms) of all living European land and freshwater animals, their geographical distribution at country level (up to the Urals, excluding the Caucasus region), and some additional information. The Fauna Europaea project covers about 230,000 taxonomic names, including 130,000 accepted species and 14,000 accepted subspecies, which is much more than the originally projected number of 100,000 species. This represents a huge effort by more than 400 contributing specialists throughout Europe and is a unique (standard) reference suitable for many users in science, government, industry, nature conservation and education.
This paper provides updated information on the taxonomic composition and distribution of the Annelida - terrestrial Oligochaeta (Megadrili and Enchytraeidae), Aphanoneura and Polychaeta, recorded in Europe. Data on 18 families, 11 autochthonous and 7 allochthonous, represented in our continent by a total of 800 species, are reviewed, beginning from their distinctness, phylogenetic status, diversity and global distribution, and following with major recent developments in taxonomic and faunistic research in Europe. A rich list of relevant references is appended. The Fauna Europaea Annelida - terrestrial Oligochaeta data-set, as completed in 2004, will be updated accordingly.
Keywords
Biodiversity Informatics, Fauna Europaea, Taxonomic indexing, Zoology, Biodiversity, Taxonomy, Annelida, Oligochaeta, terrestrial, Megadrili, Enchytraeidae, Propappidae, Aphanoneura , non-marine Polychaeta
Introduction
The European Commission published the European Community Biodiversity Strategy, providing a framework for development of Community policies and instruments in order to comply with the Convention on Biological Diversity. This Strategy recognises the current incomplete state of knowledge at all levels concerning biodiversity, which is a constraint on the successful implementation of the Convention. Fauna Europaea contributes to this Strategy by supporting one of the main themes: to identify and catalogue the components of European biodiversity into a database in order to serve as a basic tool for science and conservation policies.
With regard to biodiversity in Europe, both science and policies depend on a knowledge of its components. The assessment of biodiversity, monitoring changes, sustainable exploitation of biodiversity, and much legislative work depend upon a validated overview of taxonomic biodiversity. Towards this end Fauna Europaea plays a major role, providing a web-based information infrastructure with an index of scientific names (including important synonyms) of all living European land and freshwater animals, their geographical distribution at country level and some additional useful information. In this sense, the Fauna Europaea database provides a unique reference for many user-groups such as scientists, governments, industries, conservation communities and educational programs.
Fauna Europaea started in 2000 as an EC-FP5 four-years project, delivering its first release in 2004. After Fifteen years of steady progress, in order to efficiently disseminate the Fauna Europaea results and to increase the acknowledgement of the Fauna Europaea contributors, novel e-Publishing tools have been applied to prepare data-papers of all major taxonomic groups. For this purpose a special Biodiversity Data Journal Series has been compiled, called Contributions on Fauna Europaea. This work was initiated during the ViBRANT project and is further supported by the recently started EU BON project. This paper holds the first publication of the Fauna Europaea Annelida-Terrestrial Oligochaeta (Enchytraeidae and Megadrili), Aphanoneura and Polychaeta data sector as a BDJ data paper.
Within the EU BON project also further steps will be made to implement Fauna Europaea as a basic tool and standard reference for biodiversity research and to evaluate taxonomic expertise capacity in Europe. The Fauna Europaea data-papers will contribute to a quality assessement on biodiversity data by providing estimates on gaps in taxonomic information and knowledge.
General description
The Fauna Europaea is a database of the scientific names and distribution of all living, currently known multicellular European land and fresh-water animal species assembled by a large network of experts, using advanced electronic tools for data collations and validation routines. An extended description of the Fauna Europaea project backgrounds can be found in
The Annelida-terrestrial Oligochaeta (Megadrili and Enchytraeidae), Aphanoneura and Polychaeta is one of the 58 Fauna Europaea major taxonomic groups. In the first release of the FaEu database (2004) it covered 735 species.
Introduction to Annelida-terrestrial Oligochaeta, Aphanoneura and Polychaeta.
Part 1: Aphanoneura and Polychaeta
Class APHANONEURA
Two families whose sister-group relationship is supported by morphological and molecular data (
Family Aeolosomatidae
Cosmopolitan family of minute worms living interstitially or epibenthically mostly in freshwater habitats. Some also occur in damp soils and forest litter; others (Hystricosoma, Aeolosoma spp.) live epizoically on freshwater crayfish; one species is marine. Represented in Europe by 3 genera (Aeolosoma, Hystricosoma, Rheomorpha) and about 25 species. Most Aeolosoma species are recognizable by possessing scattered epithelial “oil glands” that are red, green or yellow in vivo, dorsal and ventral bundles of capillary chaetae, and a ciliated prostomium used for locomotion and suction-feeding. Hystricosoma moves by muscular contractions of the body wall, has only sigmoid chaetae and those of each dorsal bundle emerge as two parallel rows running in opposite directions; the orange-red oil glands are concentrated around the dorsal chaetal bundles. Rheomorpha has pale greenish oil glands and bears adhesive epidermal papillae in place of the chaetae and at the tip of the bilobed pygidium. Testes develop in both anterior and posterior segments; mature sperm are released through the nephridia and transferred into 2-5 pairs of simple ventral epidermal invaginations of the partner. Eggs are laid inside cocoons secreted by a glandularized epithelium that extends for one segment laterally and ventrally beneath the single mature ovary; the unpaired female pore opens in the middle of this pseudo-clitellum. Sexual reproduction, however, is rarely observed, reproduction most often implies paratomic formation of chains of 2-8 zooids.
Family Potamodrilidae
Monotypic family, with one species in Europe (Potamodrilus fluviatilis) and one undescribed congener (Potamodrilus sp.) in North America (
Class POLYCHAETA
Family Nerillidae
A family of minute worms endowed with small cephalic palps and antennae and cirri along the body and on the pygidium. They comprise 17 genera and about 48 species, mostly marine interstitial, occurring worldwide from the intertidal to abyssal depths (
Family Parergodrilidae
This family of unclear phylogenetic position comprises only two species: the marine littoral mesopsammic Stygocapitella subterranea and the truly terrestrial Parergodrilus heideri (
Family Serpulidae
A worldwide-distributed family of polychaetes building tubes of calcium carbonate, comprising about 350 species, nearly all marine. Marifugia cavatica Absolon & Hrabě, 1930 is the sole member living in fresh water and is a stygobiont, endemic to the European alpine-dinaric karst. Molecular phylogenetics places M. cavatica as sister taxon to a clade of brackish-water Ficopotamus species, suggesting that the transition to the subterranean environment occurred via ancestral marine shallow water to intertidal or estuarine species (
Polychaeta incertae sedis
Hrabeiella periglandulata Pižl & Chalupský, 1984 is, along with the parergodrilid Parergodrilus heideri Reisinger, 1925, the sole truly terrestrial non-clitellate annelid (
Part 2: Oligochaeta
Class OLIGOCHAETA
Family Acanthodrilidae
Allochthonous megadrile family with a very wide geographic distribution, most probably non monophyletic. Endemic taxa can be found throughout the southern continents, plus North and Central America (
Family Ailoscolecidae
Monotypic megadrile family endemic to southwestern France. They were regarded as possibly close to the North American Komarekionidae (
Family Criodrilidae
Monotypic megadrile family indigenous to the western Palaearctic. Primitively aquatic, Criodrilus lacuum Hoffmeister, 1845 is characterized by a quadrangular body section, the lack of gizzards and other gut specializations, the long clitellum, absence of spermathecae and tubercula pubertatis, and the production of long, horn-shaped spermatophores and long cocoons. Molecular data place it as sister taxon to the Hormogastridae and Lumbricidae (
Family Enchytraeidae
Microdrile family with worldwide distribution, Polar regions included. The Enchytraeidae are well separated from all other oligochaetes by a set of somatic (chaetae short and simple-pointed, emerging from a generally smooth body wall; prostomium pierced by a coelomic pore; pharyngeal glands as conspicuous masses arranged pairwise in segments 4-6, with ventral strands of ducts directed to the roof of pharynx; holonephridia with postseptal loops more or less anastomosed to form canalized compact bodies; coelomocytes abundant) and reproductive features (spermathecae opening in segment 5; testes in 11, ovaries in 12, with respective paired pores in 12 and 13; preseptal sperm funnel glandular, opposite end of vas deferens generally surrounded by a copulatory cushion). So far, the more inclusive molecular phylogenetic analyses (e.g.
The vernacular name of the family, pot-worms (fr. Greek 'en-chytra' = in flowerpot), does not reflect the great ecological diversity of the group, which, although most represented in terrestrial soils (Fig.

About half of the currently accepted inland species have their type localities in Europe, where collecting and taxonomic work has always been comparatively intense. The northernmost latitudes of the Holarctic, particularly Beringia (refs. in
The correct evaluation of species taxonomic status and distribution is a fundamental prerequisite in assessing biodiversity in any geographical region. Both these aspects are still in a state of uncertainty for many European components of the family. According to the published records, a fair number of species would appear Holarctic or even cosmopolitan. However, besides the mentioned sampling biases, many records date back to a time when identifications were based on either ambiguous diagnoses and/or inadequate examinations. Since the mid-1990s, there has been an effort to better characterize the species through an enlarged and univocal set of features (e.g.
Family Eudrilidae
The Eudrilidae, a megadrile family of tropical West and East Africa, are separated from all other earthworms by their specialized “spermathecal systems”, which are found posterior to the testis segments and are connected to the oviducts. One species, Eudrilus eugeniae, known in the fish bait market as the “African night crawler”, is a native to West Africa that has become pantropical at low altitudes. It is a relatively large earthworm that under constant high temperatures (25°–30°C) grows extremely rapidly, is prolific and thus exploited for protein production, composting and agriculture. Its main disadvantages are sensitivity to handling and its narrow temperature tolerance (it grows very slowly at 15°C and dies below 5°C;
Family Glossoscolecidae
Megadrile earthworms indigenous to the tropical forests of South and Central America, with clitellum beginning near segment 14, endowed with tubercula pubertatis. Male pores either inconspicuous or within copulatory chambers. Dorsal pores lacking, oesophageal gizzard in segment 6; extramural calciferous glands in some or all of segments 7-14; typhlosole present. Holoic with nephridial bladders in intestinal region. Spermathecae adiverticulate, in front of the gonadal segments. The family in this classical acception harbours 200 species and 25 genera, but molecular phylogeny has recently revealed it as polyphyletic (
Family Hormogastridae
Megadrile family endemic to the western Mediterranean region, whose distinctness from the common European earthworms (Lumbricidae) was first recognized by Francesco Redi (
Family Lumbricidae
Megadrile family endemic to the Holarctic (Figs


The Lumbricidae are most closely related to the Hormogastridae, Ailoscolecidae, Lutodrilidae and Criodrilidae, all families endemic to our continent (
In the first version of the Fauna Europaea database in 2004, the family Lumbricidae was represented by 384 species classified in 32 genera and the three subfamilies mentioned above, following criteria that took into account classical views and well-grounded modern morpho-taxonomy. Molecular cladistic analyses focusing on earthworms had then just started and for about a decade phylogenetic reconstructions of the Lumbricidae would be hindered by restricted taxonomic sampling and/or the low signal of the chosen genes (
At the higher levels, while the molecular data in
Family Megascolecidae
Allochthonous megadrile family, very widely distributed, whose most ancient taxa show a predominant southeastern distribution (Australia, New Zealand, southeastern Asia), but with a tribe, the Argilophilini, endemic to western North America (
Family Ocnerodrilidae
Allochthonous family of small, filiform, semiaquatic megadriles, sister taxon to the Megascolecidae s.l. (
Family Octochaetidae
The earthworm genus Dichogaster is a large, heterogeneous taxon comprising some 350 species allocated to three subgenera (
Endemic species of Dichogaster are restricted to Central and South America and Africa, but some congeners have spread synanthropically in most parts of the world. Particularly two of them, D. bolaui and D. saliens, are among the most frequent peregrine species in tropical and subtropical regions. The prominent vascularization of their body wall and inner organs facilitates their survival and spreading under flooded conditions. According to
Family Propappidae
Monogeneric microdrile family, endemic to the Palaearctic region. Although fully aquatic, it is dealt with in this paper because of its historical taxonomic association with the Enchytraeidae. The first Propappus species (P. glandulosus Michaelsen, 1905) was discovered in Lake Baikal and placed in the Enchytraeidae, to represent an early branch within the family. It would be later recorded also in surrounding rivers in Russia (
The only propappid species occurring in Europe, the proboscidate, interstitial P. volki, appears to be a good bioindicator for unpolluted water (
Family Sparganophilidae
Megadrile family of very slender worms, measuring as adults 70-200 mm in length and 2-3.5 mm in maximal width, spending their life in the mud or amongst the roots of aquatic plants. The family comprises one genus and about a dozen species, nearly all restricted to the southeastern USA. The type species, Sparganophilus tamesis Benham, 1892, originally discovered in River Thames, England (
Besides England, the occurrence of the family outside of America is limited to France, Switzerland, Germany, Italy (
Project description
This BDJ data paper includes and updates the taxonomic indexing efforts in the Fauna Europaea on European Annelida-Oligochaeta (terrestrial: Enchytraeidae and Megadrili), Aphanoneura and Polychaeta covering the first two versions of Fauna Europaea worked on between 2000 and 2013 (up to version 2.6).
The taxonomic framework of Fauna Europaea includes partner institutes, providing taxonomic expertise and information, and expert networks taking care of data collation.
Every taxonomic group is covered by at least one Group Coordinator responsible for the supervision and integrated input of taxonomic and distributional data of a particular group. The Fauna Europaea checklist would not have reached its current level of completion without the input from several groups of specialists. The formal responsibility of collating and delivering the data of relevant families rested with a number of Taxonomic Specialists (see Table
Responsible specialists per family in Annelida Oligochaeta (terrestrial: Enchytraeidae and Megadrili), Aphanoneura and Polychaeta. The numbers of databased species is given per family (see also Fig.
| TAXONOMY | EUROPE | ||||
| CLASS | FAMILY | SPECIALIST(S) | DATABASED SPECIES (Fauna Europaea) | TOTAL DESCRIBED SPECIES (information-gap) | COMMENT |
| Aphanoneura | Aeolosomatidae | Emilia Rota | 24 | 23 | Aeolosoma gineti transferred to the oligochaete family Parvidrilidae |
| Aphanoneura | Potamodrilidae | Emilia Rota | 1 | 1 | |
| Polychaeta | Nerillidae | Emilia Rota | 1 | 1 | |
| Polychaeta | Parergodrilidae | Emilia Rota | 2 | 2 | Including Hrabeiella periglandulata, incertae sedis species |
| Polychaeta | Serpulidae | Emilia Rota | 1 | 1 | |
| Oligochaeta | Acanthodrilidae | Emilia Rota | 2 | 2 | |
| Oligochaeta | Ailoscolecidae | Emilia Rota | 1 | 1 | |
| Oligochaeta | Criodrilidae | Emilia Rota | 2 | 2 | |
| Oligochaeta | Enchytraeidae | Emilia Rota | 233 | 274 | By addition of 2 new records, 39 new species, 6 synonymies, 6 reinstated species |
| Oligochaeta | Eudrilidae | Emilia Rota | 1 | 1 | |
| Oligochaeta | Glossoscolecidae | Emilia Rota | 1 | 1 | |
| Oligochaeta | Hormogastridae | Emilia Rota | 11 | 22 | By addition of 2 new species and 9 reinstated species |
| Oligochaeta | Lumbricidae | Emilia Rota | 432 | 446 | By addition of 17 new species and 3 synonymies |
| Oligochaeta | Megascolecidae | Emilia Rota | 13 | 14 | One new record in greenhouses |
| Oligochaeta | Ocnerodrilidae | Emilia Rota | 3 | 3 | |
| Oligochaeta | Octochaetidae | Emilia Rota | 4 | 4 | |
| Oligochaeta | Propappidae | Emilia Rota | 1 | 1 | |
| Oligochaeta | Sparganophilidae | Emilia Rota | 2 | 1 | By deletion of 1 synonym |
Data management tasks are taken care of by the Fauna Europaea project bureau. During the project phase (until 2004) a network of principal partners took care about diverse management tasks: Zoological Museum Amsterdam (general management & system development), Zoological Museum of Copenhagen (data collation), National Museum of Natural History in Paris (data validation) and Museum and Institute of Zoology in Warsaw (NAS extension). Since the formal project ending (2004-2013) all tasks have been undertaken by the Zoological Museum Amsterdam. Since 2013 the data servers are hosted at the Museum für Naturkunde in Berlin (migrated from ZMA-UvA).
On the available expert capacity, presently, in Europe faunistic, systematic and taxonomic studies on terrestrial Annelida (clitellate and non-clitellate) species are actively carried out in Italy (University of Siena), Hungary (Eötvös Loránd University; Eszterházy Károly College), Germany (IFAB Institut für Angewandte Bodenbiologie; ECT Oekotoxikologie; Senckenberg Research Center for Biodiversity and Climate), Denmark (University of Copenhagen), Spain (Universidad Complutense de Madrid, Universidade da Coruña, University of Vigo), Romania (Institute of Biological Research), Czech Republic (Masaryk University), Poland (Polish Academy of Sciences), UK (Natural History Museum, London; University of Cardiff), Sweden (University of Göteborg) and in Ireland (University College Dublin), by around 20 specialists, often with expertise in one or two familes. Megadriles and Enchytraeidae are generally investigated by separate communities, the one involved in enchytraeid alpha-taxonomy being composed of around six experts in Europe (and world-wide). Additional ultrastructural research is carried out in Poland (University of Silesia) and Germany (University of Osnabrück). Outside Europe around ten more specialists contribute to the taxonomy of terrestrial (clitellate and non-clitellate) Annelida, and phylogenetic studies are presently mostly conducted as intercontinental collaborations (often with North American laboratories: University of Guelph, Canada; University of Iowa; University of Washington; Harvard University; US National Museum of Natural History, Smithsonian Institution).
The area study covers the European mainland (Western Palearctic), including the Macaronesian islands, excluding the Caucasus, Turkey, Arabian Peninsula and Northern Africa (see: Geographic coverage section).
Standards. Group coordinators and taxonomic specialists have to deliver the (sub)species names according to strict standards. The names provided by Fauna Europaea are scientific names. The taxonomic scope includes issues like, (1) the definition of criteria used to identify the accepted species-group taxa, (2) the hierarchy (classification scheme) for the accommodation of all the accepted species and (3), relevant synonyms, and (4) the correct nomenclature. The Fauna Europaea 'Guidelines for Group Coordinators and Taxonomic Specialists', include the standards, protocols, scope, and limits that provide the instructions for more than 400 specialists contributing to the project, strictly following the provisions of the current edition of the International Code of Zoological Nomenclature.
Data management. The data records could either be entered offline into a preformatted MS-Excel worksheet or directly into the Fauna Europaea transaction database using an online browser interface. The data servers were hosted at the University of Amsterdam (Amsterdam academic computing facilities) until 2013, when hosting was migrated to the Museum für Naturkunde in Berlin.
Data set. The Fauna Europaea basic data set consists of: accepted (sub)species names (including authorship), synonym names (including authorship), a taxonomic hierarchy/classification, misapplied names (including misspellings and alternative taxonomic views), homonym annotations, expert details, European distribution (at country level), Global distribution (only for European species), taxonomic reference (optional), occurrence reference (optional).
Fauna Europaea was funded by the European Commission under the Fifth Framework Programme and contributed to the Support for Research Infrastructures work programme with Thematic Priority Biodiversity (EVR1-1999-20001) for a period of four years (1 March 2000 - 1 March 2004), including a short 'NAS extension', allowing EU candidate accession countries to participate. Follow-up support was given by the EC-FP6 EDIT project (GCE 018340), by the EC-FP7 PESI project (RI-223806) and by the EC-FP7 ViBRANT project (RI-261532). Continuing management and hosting of the Fauna Europaea services was supported by the University of Amsterdam (Zoological Museum Amsterdam) and SARA/Vancis. Recently the continuation of Fauna Europaea was taken over by the Museum für Naturkunde in Berlin, supported by the EC-FP7 EU BON project (grant agreement №308454).
Sampling methods
See spatial coverage and geographic coverage descriptions.
Fauna Europaea data have been assembled by principal taxonomic experts, based on their individual expertise, including literature sources, collection research, and field observations. In total no less than 476 experts contributed taxonomic and/or faunistic information for Fauna Europaea. The vast majority of the experts are from Europe (including EU non-member states). As a unique feature, Fauna Europaea funds were set aside for rewarding/compensating for the work of taxonomic specialists and group coordinators.
To facilitate data transfer and data import, sophisticated on-line (web interfaces) and off-line (spreadsheets) data-entry routines were built, integrated within an underlying central Fauna Europaea transaction database (Fig.
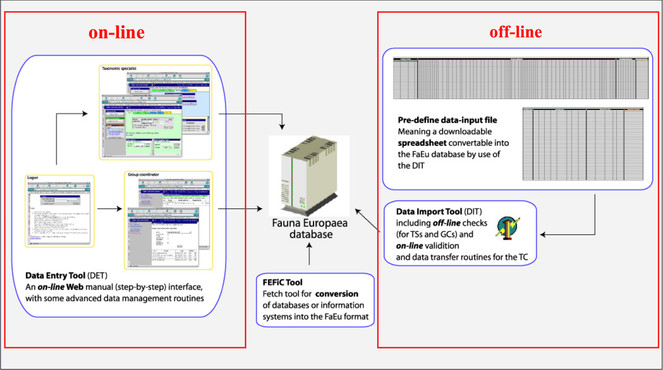
A first release of the Fauna Europaea index via the web-portal had been presented at 27th of September 2004, the most recent release (version 2.6.2) was launched on 29 August 2013. An overview of Fauna Europaea releases can be found here: http://www.faunaeur.org/about_fauna_versions.php.
Fauna Europaea data are unique in the sense that they are fully expert based. Selecting leading experts for all groups assured the systematic reliability and consistency of the Fauna Europaea data.
Furthermore, all Fauna Europaea data sets are intensively reviewed at regional and thematic validation meetings, at review sessions on taxonomic symposia (for some groups), by Fauna Europaea Focal Points (during the FaEu-NAS and PESI projects) and by various end-users sending annotations using the web form at the web-portal. Additional validation on gaps and correct spelling is being done by the validation office in Paris.
In general we expect to get taxonomic data for 99.3% of the known European fauna after the initial release. The faunistic coverage is not quite as good, but is nevertheless 90-95% of the total fauna. For terrestrial Annelida-Oligochaeta (terrestrial) the current taxonomic coverage is about 93% (see Table
Gaps of knowledge in this group are difficult to quantify. For some families, the number of existing species has remained unchanged for years and only molecular taxonomy may alter it. With regard to the Enchytraeidae, Lumbricidae and Hormogastridae, not only barcoding analyses of common taxa continuously uncover distinct lineages which may represent cryptic species, but investigations conducted in insufficiently explored areas of our continent do not cease to provide evidence of new morphospecies.
Checks on technical and logical correctness of the data have been implemented in the data entry tools, including around 50 business rules. This validation tool proved to be of huge value for both the experts and project management, and contributed significantly to preparation of a remarkably clean and consistent data set. This thorough reviewing makes Fauna Europaea the most scrutinised data set in its domain.
To optimise the use and implementation of a uniform and correct nomenclature, a cross-referencing of the Fauna Europaea Annelida-Oligochaeta (terrestrial: Enchytraeidae and Megadrili), Aphanoneura and Polychaeta data-set with relevant nomenclators, including Nomenclatura Oligochaetologica, compiled by John W. Reynolds and Mark J. Wetzel, is recommended, following the global efforts on establishing 'ZooBank' as a component of a so-called 'Global Names Architecture' (
By evaluating team structure and life cycle procedures (data-entry, validation, updating, etc.), clear definitions of roles of users and user-groups, according to the taxonomic framework were established, including ownership and read and write privileges, and their changes during the project's life-cycle. In addition, guidelines on common data exchange formats and codes have been issued (see also the 'Guidelines for Experts' document).
Geographic coverage
Species and subspecies distributions in Fauna Europaea are registered at least to country level, i.e. for political countries. For this purpose the FaEu geographical system basically follows the TDWG standards. The covered area includes the European mainland (Western Palearctic), plus the Macaronesian islands (excl. Cape Verde Islands), Cyprus, Franz Josef Land and Novaya Zemlya. Western Kazakhstan and the Caucasus are excluded (see Fig.
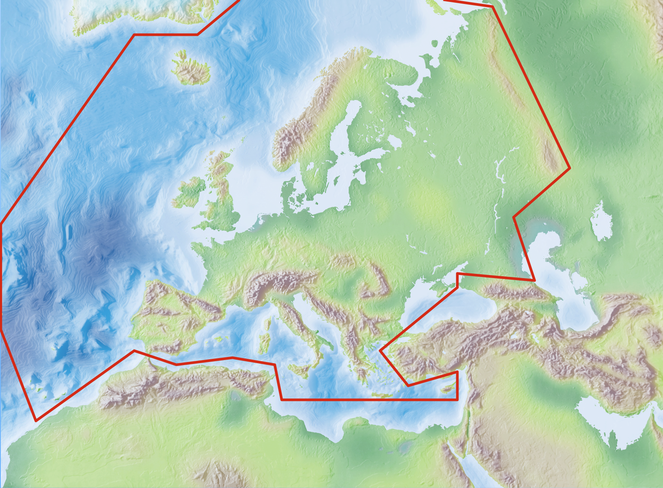
The focus is on species (or subspecies) of European multicellular animals of terrestrial and freshwater environments. Species in brackish waters, occupying the marine/freshwater or marine/terrestrial transition zones, are generally excluded.
Additional notes and updating information on the geographic coverage of Annelida – Oligochaeta (terrestrial: Enchytraeidae and Megadrili), Aphanoneura and Polychaeta in Fauna Europaea can be found below:
Family Aeolosomatidae
On global coverage:
On European coverage: New records for Portugal includes (
Family Potamodrilidae
On global coverage: Potamodrilus sp. is observed in North America (
On European coverage: A new record for Turkey includes: P. fluviatilis (
Family Nerillidae
On global coverage: T. beranecki North American records: New Hampshire, Pennsylvania, Ohio (http://collections.mnh.si.edu/).
Family Parergodrilidae
On European and global coverage: Parergodrilus heideri new records includes: Denmark (
Polychaeta incertae sedis
On European and global coverage: Hrabeiella periglandulata new records: Austria, Spain, Denmark, Hungary, Romania (
Family Acanthodrilidae
On global coverage: new records: Japan (EPA38) (
Family Enchytraeidae
In the last 15 years, many new and known species of enchytraeids have been described in good detail. This effort and the sampling of new areas and environments have not only increased the length of the local inventories (for some countries the list has been almost doubled) but also improved their quality.
First European records include: Enchytraeus luxuriosus Schmelz & Collado, 1999 and Hemifridericia bivesiculata Christensen & Dózsa-Farkas, 2006. New regional records (since 2004) fall into all geographical units and are too substantial to detail here.
Family Eudrilidae
On European coverage: Eudrilus eugeniae new records: Hungary (
Family Glossoscolecidae
On European coverage: Pontoscolex corethrurus new records: erroneously omitted for England GB-GRB (greenhouses at Kew Gardens;
Family Lumbricidae
In addition to the newly discovered species, in the last 10 years single new records, checklists and biogeographical analyses have been produced for the lumbricid faunas of several countries (e.g. Portugal, Ireland, Great Britain, Germany, Hungary, Romania, Bulgaria, Croatia, Bosnia Herzegovina, Serbia, Montenegro, Macedonia, Albania, Greece) and mountain systems (Carpathians, Stara Planina, Balkans) (
Family Megascolecidae
On European coverage: Pithemera bicincta new records: greenhouses in Hungary (
Family Ocnerodrilidae
On European coverage: Eukerria saltensis new records: France (Corsica) FR-COR, England GB-GRB (
Family Octochaetidae
On European coverage: Dichogaster bolaui new records: Ireland (
Family Propappidae
On global coverage: Propappus volki new record: Japan (
Family Sparganophilidae
On European coverage: Sparganophilus tamesis new records: Germany (
Mediterranean (N 35°) and Arctic Islands (N 82°) Latitude; Atlantic Ocean (Mid-Atlantic Ridge) (W 30°) and Ural (E 60°) Longitude.
Taxonomic coverage
The Fauna Europaea database contains the scientific names of all living European land and freshwater animal species, including numerous infra-groups and synonyms. More details about the conceptual background of Fauna Europaea and standards followed are described above and in the project description paper(s).
This data paper covers the Annelida Oligochaeta (terrestrial: Enchytraeidae and Megadrili), Aphanoneura and Polychaeta content of Fauna Europaea, including 18 families, updated from 735 species and 43 (sub)species synonyms (see Fig.
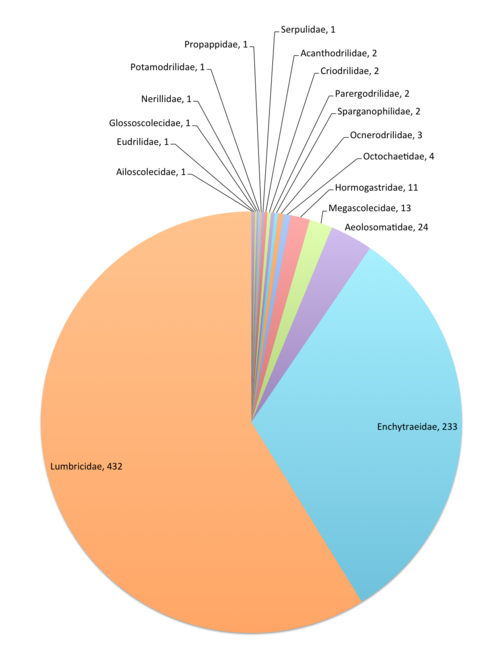
Additional notes and details of updating information on the taxonomic status and coverage of Annelida – Oligochaeta (terrestrial), Aphanoneura and Polychaeta in Fauna Europaea can be found below:
Family Aeolosomatidae
According to
The correct year of publication of A. tenebrarum by Vejdovský is 1882.
Family Enchytraeidae
1. Taxonomic novelties
A useful key to the European terrestrial enchytraeids has recently been published by
The taxonomic novelties considered here are the following:
1.1. Newly described genera
Globulidrilus Christensen & Dózsa-Farkas, 2012
1.2. Newly described species
Achaeta antefolliculata Dózsa-Farkas & Boros, 2005
Achaeta borbonica Rota, 2015
Achaeta coimbrensis Schmelz & Collado, 2013
Achaeta diddeni Graefe, 2007
Achaeta giustii Rota, 2015
Achaeta unibulba Graefe, Dózsa-Farkas & Christensen, 2009
Cernosvitoviella longiducta Dumnicka, 2010
Cernosvitoviella tridentina Dumnicka, 2004
Chamaedrilus chalupskyi Martinsson, Rota & Erséus, 2014
Chamaedrilus pseudosphagnetorum Martinsson, Rota & Erséus, 2014
Chamaedrilus varisetosus Martinsson, Rota & Erséus, 2015
Cognettia valeriae Dumnicka, 2010
Fridericia argillae Schmelz, 2003
Fridericia bargaglii Rota, 2015
Fridericia brunensis Schlaghamerský, 2007
Fridericia ciliotheca Schmelz & Collado, 2013
Fridericia crassiductata Dózsa-Farkas & Cech, 2006
Fridericia cusanica Schmelz, 2003
Fridericia dozsae Schmelz, 2003
Fridericia granosa Schmelz, 2003
Fridericia gyromonodactyla Boros & Dózsa-Farkas, 2015
Fridericia healyae Schmelz, 2003 (pro F. polychaeta Bretscher, 1900 sensu
Fridericia lacii Dózsa-Farkas, 2009
Fridericia larix Schmelz & Collado, 2005
Fridericia longeaurita Boros & Dózsa-Farkas, 2015
Fridericia lenta Schmelz, 2003 (pro F. leydigii (Vejdovský, 1878) sensu
Fridericia mahunkai Dózsa-Farkas, 2013
Fridericia marginata Schmelz & Collado, 2013
Fridericia meridiana Rota, 2015
Fridericia rara Rota, 2015
Fridericia roembkei Schmelz & Collado, 2013
Fridericia schmelzi Cech & Dózsa-Farkas, 2005
Fridericia sousai Schmelz & Collado, 2013
Fridericia transylvanica Boros & Dózsa-Farkas, 2015
Marionina deminuta Rota, 2013
Marionina mendax Rota, 2013
Marionina mimula Rota, 2013
Marionina scintillans Boros & Dózsa-Farkas, 2008
Marionina sexdiverticulata Dózsa-Farkas, 2002
1.3. New names and synonymies
Bryodrilus librus (Nielsen & Christensen, 1959) <= B. parvus Nurminen, 1970 jun. syn.
Cernosvitoviella aggtelekiensis Dózsa-Farkas, 1970 <= C. goodhui Healy, 1975 jun. syn.
Cernosvitoviella palustris Healy, 1979 <= C. estaragniensis Giani, 1979 jun. syn.
Enchytraeus dichaetus Schmelz & Collado, 2010 nom. nov. pro E. minutus bisetosus Rota & Healy, 1994
Fridericia minor Friend, 1913 <= F. gracilis von Bulow, 1957 jun. syn.
Fridericia miraflores Sesma & Dózsa-Farkas, 1996 <= F. sylvatica Healy, 1979 nom. dub.
Mesenchytraeus pelicensis Issel, 1905 <= M. kuehnelti Dózsa-Farkas, 1991 jun. syn.
Fridericia Michaelsen, 1889 <= Timmodrilus Dózsa-Farkas, 1997 jun. syn.
1.4. New combinations
Bryodrilus librus (Nielsen & Christensen, 1959)
Chamaedrilus anomalus (Černosvitov, 1928)
Chamaedrilus cognettii (Issel, 1905)
Chamaedrilus glandulosus (Michaelsen, 1888)
Chamaedrilus hibernicus (Healy, 1975)
Chamaedrilus lapponicus (Nurminen, 1965)
Chamaedrilus paxi (Moszyński, 1938)
Chamaedrilus sphagnetorum (Vejdovský, 1878)
Chamaedrilus valeriae (Dumnicka, 2010)
Euenchytraeus clarae (Bauer, 1993)
Globulidrilus riparius (Bretscher, 1899)
1.5. Reinstatements as valid names
Chamaedrilus Friend, 1913 pro Cognettia Nielsen & Christensen, 1959 (partim)
Euenchytraeus Bretscher, 1906 pro Cognettia Nielsen & Christensen, 1959 (partim)
Buchholzia subterranea (Černosvitov, 1937)
Chamaedrilus chlorophilus Friend, 1913
Enchytraeus bohemicus Dumnicka, 1996
Fridericia digitata Cognetti, 1901
Fridericia glandifera Friend, 1913
Fridericia humicola Bretscher, 1900
1.6. Species complexes
The following names are considered as species complexes, their taxonomy being currently still unresolved:
Enchytraeus buchholzi s.l.
Enchytronia parva s.l.
Fridericia aurita s.l.
Fridericia ratzeli s.l.
1.7. Rejected synonymies
Achaeta etrusca Rota, 1995 ≠ A. iberica Graefe, 1989
Fridericia caprensis Bell, 1947 ≠ F. pretoriana Stephenson, 1930
Fridericia sohlenii Rota, Healy & Erséus, 1998 ≠ F. cylindrica Springett, 1971
Fridericia glandulosa Southern, 1907 ≠ F. galba (Hoffmeister, 1843)
Family Hormogastridae
The correct year of publication of Vignysa Bouché is 1970; same for Vignysa popi Bouché, 1970 (both recorded as 1972 in the 2004 release).
Taxonomic comments: H. elisae species complex comprises at least five cryptic allopatric species.
Species to be added as valid: H. riojana, H. ireguana, H. eserana, H. huescana, H. arenicola, H. catalaunensis, H. sylvestris, H. najaformis, H. castillana (no H. multilamella, H. lleidana), all authored 'Qiu & Bouché, 1998'
Hormogaster abbatissae Novo et al., 2012a
Hormogaster joseantonioi Fernández Marchán et al., 2014
Family Lumbricidae
1. Taxonomic novelties
Since the first version of the Fauna Europaea database in 2004, some 17 new species have been described from our continent and the status of some known species has been formally revised (
1.1. Newly described species:
Allolobophora prosellodacica Csuzdi & Pop, 2008 (herein transferred to Eophila)
Allolobophora ruzsai Szederjesi, 2014
Dendrobaena luraensis Szederjesi & Csuzdi, 2012
Dendrobaena retrosella Szederjesi & Csuzdi, 2012
Dendrobaena virgata Szederjesi et al., 2014
Dendrobaena vladeasa Csuzdi et al., 2011
Eisenia muranyii Szederjesi & Csuzdi, 2015
Eisenia oreophila Szederjesi & Csuzdi, 2012
Eiseniona gerardoi Díaz-Cosín et al., 2014
Eumenescolex proclitellatus Perez-Onteniente & Rodriguez-Babio, 2004
Kenleenus armadas Blakemore, 2012 (pro Prosellodrilus amplisetosus Bouché, 1972 sensu
Octodrilus albanicus Szederjesi & Csuzdi, 2015
Octodrilus izanus Csuzdi et al., 2011
Octodrilus juvyi Zicsi & Cuendet, 2005
Octodrilus parvivesiculatus Csuzdi et al., 2011
Proctodrilus thaleri Höser & Zicsi, 2009
Zophoscolex albacetensis Perez-Onteniente & Rodriguez-Babio, 2010 (herein transferred to Aporrectodea)
1.2. New synonymies:
Eophila getica (Pop, 1947) <= Allolobophora dugesi var. getica = Cernosvitovia (Zicsiona) getica Mršić & Šapkarev, 1987 = Cernosvitovia (Zicsiona) silicata Mršić & Šapkarev, 1987 = Cernosvitovia (Zicsiona) paradoxa Mršić, 1992; revision by
Dendrobaena attemsi Michaelsen, 1902 <= Dendrobaena apora Qiu & Bouché, 1998; revision by
Octolasion cyaneum (Savigny, 1826) <= Dendrobaena jeanneli Pop, 1948 revision by
1.3. New combinations
Aporrectodea albacetensis (Perez-Onteniente & Rodriguez-Babio, 2010)
Eophila prosellodacica (Csuzdi & Pop, 2008)
Prosellodrilus armadas (Blakemore, 2012)
Family Sparganophilidae
Sparganophilus langi Qiu & Bouché, 1998 from Switzerland is a junior synonym of S. tamesis Benham, 1892 (
2. Classification
The oligochaete suprafamilial rankings, as they were published in FaEu 2004 and listed below, need revision. No adjustments to the rankings were introduced in this paper.
| Rank | Scientific Name |
|---|---|
| kingdom | Animalia |
| subkingdom | Eumetazoa |
| phylum | Annelida |
| class | Aphanoneura |
| family | Aeolosomatidae |
| family | Potamodrilidae |
| class | Oligochaeta |
| subclass | Diplotesticulata |
| superorder | Megadrili |
| order | Opisthopora |
| suborder | Lumbricina |
| superfamily | Criodriloidea |
| family | Criodrilidae |
| superfamily | Eudriloidea |
| family | Eudrilidae |
| superfamily | Lumbricoidea |
| family | Ailoscolecidae |
| family | Glossoscolecidae |
| family | Hormogastridae |
| subfamily | Hormogastrinae |
| subfamily | Vignysinae |
| subfamily | Xaninae |
| family | Lumbricidae |
| subfamily | Diporodrilinae |
| subfamily | Lumbricinae |
| subfamily | Spermophorodrilinae |
| superfamily | Megascolecoidea |
| family | Acanthodrilidae |
| family | Megascolecidae |
| family | Ocnerodrilidae |
| family | Octochaetidae |
| superfamily | Sparganophiloidea |
| family | Sparganophilidae |
| subclass | Tubificata |
| order | Tubificida |
| suborder | Enchytraeina |
| superfamily | Enchytraeoidea |
| family | Enchytraeidae |
| family | Propappidae |
| class | Polychaeta |
| order | Nerillida |
| family | Nerillidae |
| order | Sabellida |
| family | Serpulidae |
| subclass | Polychaeta incertae sedis |
| family | Parergodrilidae |
Temporal coverage
Currently living animals in stable populations, largely excluding (1) rare/irregular immigrants, intruder or invader species, (2) accidental or deliberate releases of exotic (pet) species, (3) domesticated animals, (4) foreign species imported and released for bio-control or (5) foreign species largely confined to hothouses.
Usage rights
Fauna Europaea data are licensed under CC BY SA version 4.0. The property rights of experts over their data is covered by their Fauna Europaea contract agreements. For more IPR details see: http://www.faunaeur.org/copyright.php.
Data resources
| Column label | Column description |
|---|---|
| datasetName | The name identifying the data set from which the record was derived (http://rs.tdwg.org/dwc/terms/datasetName). |
| version | Release version of data set. |
| versionIssued | Issue data of data set version. |
| rights | Information about rights held in and over the resource (http://purl.org/dc/terms/rights). |
| rightsHolder | A person or organization owning or managing rights over the resource (http://purl.org/dc/terms/rightsHolder). |
| accessRights | Information about who can access the resource or an indication of its security status (http://purl.org/dc/terms/accessRights). |
| taxonID | An identifier for the set of taxon information (http://rs.tdwg.org/dwc/terms/taxonID) |
| parentNameUsageID | An identifier for the name usage of the direct parent taxon (in a classification) of the most specific element of the scientificName (http://rs.tdwg.org/dwc/terms/parentNameUsageID). |
| scientificName | The full scientific name, with authorship and date information if known (http://rs.tdwg.org/dwc/terms/scientificName). |
| acceptedNameUsage | The full name, with authorship and date information if known, of the currently valid (zoological) taxon (http://rs.tdwg.org/dwc/terms/acceptedNameUsage). |
| originalNameUsage | The original combination (genus and species group names), as firstly established under the rules of the associated nomenclaturalCode (http://rs.tdwg.org/dwc/terms/originalNameUsage). |
| family | The full scientific name of the family in which the taxon is classified (http://rs.tdwg.org/dwc/terms/family). |
| familyNameId | An identifier for the family name. |
| genus | The full scientific name of the genus in which the taxon is classified (http://rs.tdwg.org/dwc/terms/genus). |
| subgenus | The full scientific name of the subgenus in which the taxon is classified. Values include the genus to avoid homonym confusion (http://rs.tdwg.org/dwc/terms/subgenus). |
| specificEpithet | The name of the first or species epithet of the scientificName (http://rs.tdwg.org/dwc/terms/specificEpithet). |
| infraspecificEpithet | The name of the lowest or terminal infraspecific epithet of the scientificName, excluding any rank designation (http://rs.tdwg.org/dwc/terms/infraspecificEpithet). |
| taxonRank | The taxonomic rank of the most specific name in the scientificName (http://rs.tdwg.org/dwc/terms/infraspecificEpithet). |
| scientificNameAuthorship | The authorship information for the scientificName formatted according to the conventions of the applicable nomenclaturalCode (http://rs.tdwg.org/dwc/terms/scientificNameAuthorship). |
| authorName | The four-digit year in which the scientificName was published (http://rs.tdwg.org/dwc/terms/namePublishedInYear). |
| namePublishedInYear | The four-digit year in which the scientificName was published (http://rs.tdwg.org/dwc/terms/namePublishedInYear). |
| Brackets | Annotation if authorship should be put between parentheses. |
| nomenclaturalCode | The nomenclatural code under which the scientificName is constructed (http://rs.tdwg.org/dwc/terms/nomenclaturalCode). |
| taxonomicStatus | The status of the use of the scientificName as a label for a taxon (http://rs.tdwg.org/dwc/terms/taxonomicStatus). |
| resourceDescription | An account of the resource, including a data-paper DOI (http://purl.org/dc/terms/description) |
| Column label | Column description |
|---|---|
| datasetName | The name identifying the data set from which the record was derived (http://rs.tdwg.org/dwc/terms/datasetName). |
| version | Release version of data set. |
| versionIssued | Issue data of data set version. |
| rights | Information about rights held in and over the resource (http://purl.org/dc/terms/rights). |
| rightsHolder | A person or organization owning or managing rights over the resource (http://purl.org/dc/terms/rightsHolder). |
| accessRights | Information about who can access the resource or an indication of its security status (http://purl.org/dc/terms/accessRights). |
| taxonName | The full scientific name of the higher-level taxon |
| scientificNameAuthorship | The authorship information for the scientificName formatted according to the conventions of the applicable nomenclaturalCode (http://rs.tdwg.org/dwc/terms/scientificNameAuthorship). |
| taxonRank | The taxonomic rank of the most specific name in the scientificName (http://rs.tdwg.org/dwc/terms/infraspecificEpithet). |
| taxonID | An identifier for the set of taxon information (http://rs.tdwg.org/dwc/terms/taxonID) |
| parentNameUsageID | An identifier for the name usage of the direct parent taxon (in a classification) of the most specific element of the scientificName (http://rs.tdwg.org/dwc/terms/parentNameUsageID). |
| resourceDescription | An account of the resource, including a data-paper DOI (http://purl.org/dc/terms/description) |
References
- New records of twelve species of Oligochaeta (Naididae and Aeolosomatidae) from the Southern Iraqi Marshes. Iraq.Jordan Journal of Biological Sciences5(2):105‑112.
- Oligochaeta and Aphanoneura (Annelida) fauna of the Gediz Delta (Menemen-Izmir).Turk J. Zool28:183‑197.
- On the geographical distribution of earthworms.Proc. Zool Soc London1983:733‑738.
- Annelida. Oligochaeta.Kew Bull Misc Info Addit Ser5:66‑67.
- A new English genus of aquatic Oligochaeta (Sparganophilus) belonging to the family Rhinodrilidae.Q. J. Microsc. Soc.34:155‑179.
- Proceedings of the IAA 19th Conference.Innsbruck, Austria,2012.
- Distribution, life cycle and population dynamics of the aquatic enchytraeid Propappus volki (Oligochaeta) in an English chalkstream.Ecography5(1):67‑75. https://doi.org/10.1111/j.1600-0587.1982.tb01019.x
- Blakemore RJ (2005) Whither Octochaetidae? – A review of its family status (Annelida: Oligochaeta). In: Pop VV, Pop AA (Eds) Advances in earthworm taxonomy II.Cluj Univ. Press,Cluj,63-84pp.
- Hibernian reports of a new Franco-Iberian worm (Oligochaeta: Megadrilacea: Lumbricidae).Opuscula Zoologica Budapest43(2):121‑130.
- Japanese earthworms revisited a decade on (Oligochaeta: Megadrilacea).Zoology in the Middle East58(4):15‑22. https://doi.org/10.1080/09397140.2012.10648981
- New insights into polychaete phylogeny (Annelida) inferred from 18S rDNA sequences.Molecular Phylogenetics and Evolution29(2):279‑288. https://doi.org/10.1016/s1055-7903(03)00107-6
- Remarques sur quelques Lumbricina de France et consequences de la decouverte des nouveaux taxons Vignysinae (Subfam. nov.) et Diporodrilidae (Fam. nov.).Pedobiologia10:246‑256.
- Lombriciens de France. Ecologie et systématique.Annales de Zoologie-Ecologie Animalenumero hors-série:1‑671.
- Bouché MB (1983) The establishment of earthworm communities. In: Satchell JE (Ed.) Earthworm Ecology: From Darwin to Vermiculture.Chapman and Hall,London. https://doi.org/10.1007/978-94-009-5965-1_38
- Un nouveau Sparganophilus (Annelida: Oligochaeta) d’Europe, avec considérations paléogéographiques sur les Lumbricina.Doc. Pédozool. Intégrol16:178‑180.
- Branko J, Bilandžija H, Cukrov M (2012) Distribution of the Dinaric cave-dwelling tube worm Marifugia cavatica Absolon & Hrabe, 1930 in Croatia. Poster presented at 21st Int. Conference on Subterranean Biology, Abstract book, Košice, Slovačka, 02.-07.09.2012.
- Evolutionary relationships within the Clitellata: an update.Megadrilogica5:109‑112.
- Are the sexual, somatic and genetic characters enough to solve nomenclatural problems in lumbricid taxonomy?Soil Biology and Biochemistry41(11):2257‑2271. https://doi.org/10.1016/j.soilbio.2009.07.008
- Phylogenetic analysis of New Zealand earthworms (Oligochaeta: Megascolecidae) reveals ancient clades and cryptic taxonomic diversity.Molecular Phylogenetics and Evolution58(1):85‑96. https://doi.org/10.1016/j.ympev.2010.09.024
- Zur Morphologie und Systematik der Aeolosomatidae Beddard 1895 und Potamodrilidae nov. fam (Oligochaeta).Zool. Jahrb. Syst94:187‑368.
- Ultrastructure of the spermatozoon and spermiogenesis in the interstitial annelid Potamodrilus fluviatilis.Journal of Morphology185(2):203‑216. https://doi.org/10.1002/jmor.1051850206
- Ultrastructural investigations on the spermatozoon and its genesis in Aeolosoma litorale with considerations on the phylogenetic implications for the aeolosomatidae (annelida).Journal of Ultrastructure and Molecular Structure Research95:113‑130. https://doi.org/10.1016/0889-1605(86)90035-2
- Mapping of earthworm distribution for the British Isles and Eire highlights the under-recording of an ecologically important group.Biodiversity and Conservation21:475‑485. https://doi.org/10.1007/s10531-011-0194-x
- Cech G, Dózsa-Farkas K (2005) Identification of Fridericia schmelzi sp. n. combining morphological characters and PCR-RFLP analysis. In: Pop VV, Pop AA (Eds) Advances in Earthworm Taxonomy II.Cluj University Press,Cluj-Napoca,99–118pp.
- Revision of Bryodrilus glandulosus (Dózsa-Farkas, 1990) and Mesenchytraeus kuehnelti Dózsa-Farkas, 1991 (Oligochaeta: Enchytraeidae) using morphological and molecular data.Zoologischer Anzeiger - A Journal of Comparative Zoology251(3):253‑262. https://doi.org/10.1016/j.jcz.2011.09.005
- Cech G, Csuzdi C, Marialigeti K (2005) Remarks on the molecular phylogeny of the genus Dendrobaena (sensus Pop 1941) based on the investigation of 18S rDNA sequences. In: Pop VV, Pop AA (Eds) Advances in Earthworm Taxonomy II (Annelida: Oligochaeta).Cluj University Press,Cluj-Napoca, Romania,85–98pp.
- Aquatic Oligochaeta of the U.S.S.R.Akademia Nauk S.S.S.R.,Moskva-Leningrad,411pp. [InRussian].
- System der Enchytraeiden.Bull. Ass. Russe Rech. Sci. Prague5:263‑295.
- A critique of earthworm molecular phylogenetics.Pedobiologia54:S3‑S9. https://doi.org/10.1016/j.pedobi.2011.07.015
- Invasion of terrestrial enchytraeids into two postglacial tundras: North-eastern Greenland and the Arctic Archipelago of Canada (Enchytraeidae, Oligochaeta).Polar Biology29(6):454‑466. https://doi.org/10.1007/s00300-005-0076-3
- A new genus Globulidrilus and three new enchytraeid species (Oligochaeta: Enchytraeidae) from Seoraksan National Park (Korea).Journal of Natural History46:2769‑2785. https://doi.org/10.1080/00222933.2012.737038
- Species diversity and distributions of microdrile earthworms (Annelida, Clitellata, Enchytraeidae) from South America.Zootaxa2065:51‑68.
- Redescription of the oligochaete genus Propappus, and diagnosis of the new family Propappidae (Annelida: Oligochaeta).Proceedings of the Biological Society of Washington99:417‑428.
- Speciation, genetic divergence and palaeogeography in the Hormogastridae.Soil Biology and Biochemistry24:1213‑1221. https://doi.org/10.1016/0038-0717(92)90096-g
- Oligochaeta and Aphanoneura in two Northern German hardwater lakes of different trophic state.Hydrobiologia406:143‑148. https://doi.org/10.1007/978-94-011-4207-6_14
- Über ein Vorkommen von Microscolex phosphoreus (Dugès, 1837) (Oligochaeta: Acanthodrilidae) in Ungarn.Opusc. Zool. Budapest22:63‑66.
- A catalogue of Benhamiinae species (Annelida: Oligochaeta: Acanthodrilidae).Ann. Naturhist. Mus. Wien97:99‑123.
- Revision der Unterfamilie Benhamiinae Michaelsen, 1897 (Oligochaeta: Acanthodrilidae).Mitt. Zool. Mus. (Berlin)72:347‑367. https://doi.org/10.1002/mmnd.4800720218
- Earthworm species, a searchable database.Opuscula Zoologica Budapest43(1):97‑99.
- Replacement names for Eutrigaster (Graffia) Csuzdi & Zicsi, 1991 and Dichogaster (Malawia) Csuzdi, 2010 (Oligochaeta, Acanthodrilidae).Zootaxa3811(1):149‑150. https://doi.org/10.11646/zootaxa.3811.1.11
- The earthworms (Oligochaeta) of Jordan.Zoology in the Middle East34(1):71‑78. https://doi.org/10.1080/09397140.2005.10638085
- Remarks on the species Dendrobaena jeanneli Pop, 1948 and its proposed synonymy with Octolasion cyaneum (Savigny, 1826) (Oligochaeta: Lumbricidae).Opuscula Zoologica Budapest36:103‑104.
- Redescription of Allolobophora dugesi getica Pop, 1947 and its allocation to the genus Cernosvitovia Omodeo, 1956 (Oligochaeta Lumbricidae).European Journal of Soil Biology43:S19‑S23. https://doi.org/10.1016/j.ejsobi.2007.08.005
- Taxonomic and biogeographic analysis of the Allolobophora sturanyi species group (Oligochaeta, Lumbricidae).Opuscula Zoologica Budapest37:23‑28.
- Is Dichogaster bolaui (Michaelsen, 1891) the first domicole earthworm species?European Journal of Soil Biology44(2):198‑201. https://doi.org/10.1016/j.ejsobi.2007.05.003
- The earthworm fauna of the Carpathian Basin with new records and description of three new species (Oligochaeta: Lumbricidae).Zoologischer Anzeiger - A Journal of Comparative Zoology250(1):2‑18. https://doi.org/10.1016/j.jcz.2010.10.001
- An annotated checklist of the earthworm fauna of Turkey (Oligochaeta: Lumbricidae).Zootaxa1175:1‑29.
- A new earthworm species within a controversial genus: Eiseniona gerardoi sp. n. (Annelida, Lumbricidae) - description based on morphological and molecular data.ZooKeys399:71‑87. https://doi.org/10.3897/zookeys.399.7273
- Domínguez J, Edwards CA (2011) Biology and ecology of earthworm species used for vermicomposting. In: Edwards CA, Arancon NQ, Sherman R (Eds) Vermiculture Technology.CRC Press Taylor and Francis Group,Florida,249-261pp.
- The biology and population dynamics of Eudrilus eugeniae (Kinberg) (Oligochaeta) in cattle waste solids.Pedobiologia45(4):341‑353. https://doi.org/10.1078/0031-4056-00091
- Underground evolution: New roots for the old tree of lumbricid earthworms.Molecular Phylogenetics and Evolution83:7‑19. https://doi.org/10.1016/j.ympev.2014.10.024
- Unexpected occurrence of Hemifridericia bivesiculata Christensen & Dózsa-Farkas, 2006 in Hungary, a species presumed to be endemic to Devon Island, Canada, and its comparative analysis with H. parva Nielsen & Christensen, 1959 (Enchytraeidae, Oligochaeta).Zootaxa3914(2):185‑194. https://doi.org/10.11646/zootaxa.3914.2.8
- Three new Hemienchytraeus species (Enchytraeidae, Oligochaeta, Annelida) from Korea, with first records of other enchytraeids and terrestrial polychaetes (Annelida).Zootaxa2406:29‑56.
- Hrabeiella periglandulata (Annelida: “Polychaeta”) – Do apparent differences in chaetal ultrastructure indicate the existence of several species in Europe?Acta Zoologica Academiae Scientiarum Hungaricae59:143‑156.
- New enchytraeid species (Enchytraeidae, Oligochaeta) from Korea.Zootaxa4006(1):2769‑2785. https://doi.org/10.11646/zootaxa.4006.1.9
- Are Bryodrilus parvus Nurminen, 1970 and Bryodrilus librus (Nielsen and Christensen, 1959) (Annelida: Enchytraeidae) really different species? A revision based on DNA barcodes and morphological data.Zootaxa3276:38‑50.
- First records of the allochthonous species Dichogaster saliens and D. bolaui (Oligochaeta: Octochaetidae) from Sweden.Megadrilogica6:17‑20.
- Molecular phylogeny of Enchytraeidae (Annelida, Clitellata).Molecular Phylogenetics and Evolution57(2):849‑858. https://doi.org/10.1016/j.ympev.2010.07.005
- Polychaete systematics: Past and present.Zoologica Scripta26(2):71‑138. https://doi.org/10.1111/j.1463-6409.1997.tb00411.x
- Fender WM (1995) Native earthworms of the Pacific Northwest: an ecological overview. In: Hendrix PF (Ed.) Earthworm Ecology and Biogeography in North America.CRC Press Lewis Publ.,Florida,53-66pp.
- New light into the hormogastrid riddle: morphological and molecular description of Hormogaster joseantonioi sp. n. (Annelida, Clitellata, Hormogastridae).ZooKeys414:1‑17. https://doi.org/10.3897/zookeys.414.7665
- Ovary ultrastructure and oogenesis in Propappus volki Michaelsen, 1916 (Annelida: Clitellata).Zoologischer Anzeiger - A Journal of Comparative Zoology257:110‑118. https://doi.org/10.1016/j.jcz.2015.05.006
- First record of the aquatic earthworm Sparganophilus tamesis Benham, 1892 (Clitellata, Sparganophilidae) in Germany.Proceedings 5th Int. Oligochaete Taxonomy Meeting,Beatenberg, Switzerland.25-25pp.
- Checklist of the earthworm fauna of Croatia (Oligochaeta: Lumbricidae).Zootaxa3710(1):1‑30. https://doi.org/10.11646/zootaxa.3710.1.1
- Records of Enchytraeidae (Oligochaeta) in Ireland.J. Life Sci. r. Dubl. Soc.1:39‑70.
- Immunohistochemical (cLSM) and ultrastructural analysis of the central nervous system and sense organs in Aeolosoma hemprichi (Annelida, Aeolosomatidae).Zoomorphology120(2):65‑78. https://doi.org/10.1007/s004350000022
- Hörner K, Moog O, Šporka F (1995) Polychaeta. In: Moog O (Ed.) Fauna Aquatica Austriaca, Lieferungen 1995, 2002.Wasserwirtschaftskataster, Bundesministerium für Land- und Forstwirtschaft, Umwelt und Wasserwirtschaft, Wien.
- Eine neue Proctodrilus-Art (Oligochaeta: Lumbricidae) und ihr spezifischer Ökotop.Contributions to Natural History12:651‑658.
- Molecular phylogeny of earthworms (Annelida: Crassiclitellata) based on 28S, 18S and 16S gene sequences.Invertebrate Systematics26(2):213‑229. https://doi.org/10.1071/is11012
- Re-erection of Rhinodrilidae Benham, 1890, a senior synonym of Pontoscolecidae James, 2012 (Annelida: Clitellata).Zootaxa3540:67‑68.
- Phylogeny of the Megascolecidae and Crassiclitellata (Annelida, Oligochaeta): combined versus partitioned analysis using nuclear (28S) and mitochondrial (12S, 16S) rDNA.Zoosystema24:707‑734.
- Jamieson BGM (1971) Family Glossoscolecidae. In: Brinkhurst RO, Jamieson BG (Eds) Aquatic Oligochaeta of the world.Univ. Toronto Press,Toronto,723-837pp.
- Naididae (Clitellata: Oligochaeta) and Aeolosomatidae (Polychaeta: Aphanoneura ) species associated with aquatic plants in Tigris River Baghdad, Iraq.J. Baghdad for Sci10(1):116‑125.
- Fauna Europaea – all European animal species on the web.Biodiversity Data Journal2:e4034. https://doi.org/10.3897/bdj.2.e4034
- First record of the earthworm Prosellodrilus amplisetosus (Oligochaeta: Lumbricidae) outside continental Europe.Irish Naturalists’ Journal32(1):26‑28.
- Ocorrencia de oligoquetas nos solos do Rio Grande do Sul.Biologica Leopoldensia1:137‑166.
- Evolution of the unique freshwater cave‐dwelling tube worm Marifugia cavatica (Annelida: Serpulidae).Systematics and Biodiversity7(4):389‑401. https://doi.org/10.1017/s1477200009990168
- A new genus of earthworms Omodeoia gen. nov. (Oligochaeta, Lumbricidae).Bulletin of the Georgian Academy of Sciences148(3):129‑134.
- New data on earthworms (Oligochaeta: Lumbricidae) Systematic.Proceedings of the Institute of Zoology Tbilisi20:77‑84. [InRussian].
- About validity of genus Omodeoia Kvavadze, 1993 (Oligochaeta, Lumbricidae).Proceedings of the Georgian Academy of Sciences, Biol., Ser. B.5(3):1‑7.
- Kleine dieren uit het zoete water van Suriname Verslag van een onderzoek in 1967.Zoologische Bijdragen12:3‑46.
- Adaptive strategies of Pontoscolex corethrurus (Glossoscolecidae, Oligochaeta), a peregrine geophagous earthworm of the humid tropics.Biology and Fertility of Soils5(3):188‑194. https://doi.org/10.1007/bf00256899
- Checklist of earthworms (Oligochaeta: Lumbricidae) from Germany.Zootaxa3866(2):221‑245. https://doi.org/10.11646/zootaxa.3866.2.3
- Structure and distribution of enchytraeid communities from Mountain Changbaishan in China, with emphasis on the influence of environmental variables.European Journal of Soil Biology47(4):223‑229. https://doi.org/10.1016/j.ejsobi.2011.06.002
- Spermiogenesis and seminal receptacles in Aeolosoma singulare (Annelida, Polychaeta, Aeolosomatidae).Italian Journal of Zoology70(2):123‑132. https://doi.org/10.1080/11250000309356505
- Primera cita en la peninsula Iberica del poliqueto terrestre Parergodrilus heideri Reisinger, 1925 (Polychaeta, Parergodrilidae).Graellsia65:235‑240. https://doi.org/10.3989/graellsia.2009.v65.i2.152
- The Parvidrilidae - a diversified groundwater family: description of six new species from southern Europe, and clues for its phylogenetic position within Clitellata (Annelida).Zoological Journal of the Linnean Society166(3):530‑558. https://doi.org/10.1111/j.1096-3642.2012.00857.x
- Cryptic diversity in the well-studied terrestrial worm Cognettia sphagnetorum (Clitellata: Enchytraeidae).Pedobiologia57(1):27‑35. https://doi.org/10.1016/j.pedobi.2013.09.006
- Revision of Cognettia (Clitellata, Enchytraeidae): re-establishment of Chamaedrilus and description of cryptic species in the sphagnetorum complex.Systematics and Biodiversity13(3):257‑277. https://doi.org/10.1080/14772000.2014.986555
- On the identity of Chamaedrilus glandulosus (Michaelsen, 1888) (Clitellata, Enchytraeidae), with the description of a new species.Zookeys501:1‑14. https://doi.org/10.3897/zookeys.501.9279
- The aquatic earthworm Criodrilus lacuum Hoffmeister in North America (Oligochaeta, Glossoscolecidae).The Wasmann Journal of Biology11(3):373‑378.
- Northward range extension of an endemic soil decomposer with a distinct trophic position.Biology Letters8:956‑959. https://doi.org/10.1098/rsbl.2012.0537
- On a collection of African Oligochaeta in the British Museum.Proceedings of the Zoological Society of London107:501‑528. https://doi.org/10.1111/j.1096-3642.1938.tb00010.x
- Distribution, biogeographical significance and status of Lumbricus meliboeus Rosa, 1884 (Oligochaeta, Lumbricidae) at the European scale: first findings in Serbia and in Bulgaria.North-Western Journal of Zoology9:63‑69.
- The Enchytraeidae, critical revision and taxonomy of European species.Natura Jutlandica8-9:1‑160.
- High genetic divergence of hormogastrid earthworms (Annelida, Oligochaeta) in the central Iberian Peninsula: evolutionary and demographic implications.Zoologica Scripta38(5):537‑552. https://doi.org/10.1111/j.1463-6409.2009.00389.x
- Understanding the biogeography of a group of earthworms in the Mediterranean basin—The phylogenetic puzzle of Hormogastridae (Clitellata: Oligochaeta).Molecular Phylogenetics and Evolution61:125‑135. https://doi.org/10.1016/j.ympev.2011.05.018
- Cryptic speciation of hormogastrid earthworms revealed by mitochondrial and nuclear data.Molecular Phylogenetics and Evolution56(1):507‑512. https://doi.org/10.1016/j.ympev.2010.04.010
- Compilation of morphological and molecular data, a necessity for taxonomy: The case of Hormogaster abbatissae sp. n. (Annelida, Clitellata, Hormogastridae).ZooKeys242:1‑17. https://doi.org/10.3897/zookeys.242.3996
- Appearances can be deceptive: different diversification patterns within a group of Mediterranean earthworms (Oligochaeta, Hormogastridae).Molecular Ecology21(15):3776‑3793. https://doi.org/10.1111/j.1365-294x.2012.05648.x
- Unearthing the historical biogeography of Mediterranean earthworms (Annelida: Hormogastridae).Journal of Biogeography42(4):751‑762. https://doi.org/10.1111/jbi.12447
- Sistematica e distribuzione geografica degli Hormogastrinae (Oligocheti).Archivio Botanico e Biogeografico Italiano32:159‑177.
- The earthworm fauna of Sardinia.Revue d’Ecologie et Biologie du Sol21:115‑126.
- Evolution and biogeography of megadriles (Annelida, Clitellata).Italian Journal of Zoology67:179‑201. https://doi.org/10.1080/11250000009356313
- Caractères originaux des peuplements des îles tyrrhéniennes en Oligochètes terricoles.Bulletin de la Société Zoologique de France112(1):197‑214.
- Omodeo P, Rota E (2004) Taxonomic remarks on the earthworms inhabiting the Western Alps. In: Moreno AG, Borges S (Eds) Advances in Earthworm Taxonomy.Advances in Earthworm Taxonomy.Editorial Complutense,Madrid, Spain,220–259pp.
- Earthworm diversity and land evolution in three Mediterranean districts.Proceedings of the California Academy of Sciences (Suppl. 1)59:65‑83.
- Taxonomic assessment of Lumbricidae (Oligochaeta) earthworm genera using DNA barcodes.European Journal of Soil Biology48:41‑47. https://doi.org/10.1016/j.ejsobi.2011.10.003
- Eumenescolex proclitellatus n. sp. (Oligochaeta, Lumbricidae), a new species of earthworm from the Valencian Community, Spain.Boletin de la Real Sociedad Espanola de Historia Natural Seccion Biologica99:55‑61.
- Zophoscolex albacetensis sp. nov. una nueva especie de lombriz de tierra procedente de Albacete, España (Annelida, Oligochaeta).Boletin de la Real Sociedad Espanola de Historia Natural Seccion Biologica104:29‑33.
- Oligochaeta limícolas de Portugal: nota sobre a família Aeolosomatidae.Boletim da Sociedade Portuguesa de Ciencias Naturais21:45‑50.
- Pinder A (2004) Annelida: Aphanoneura. In: Yule C, Yong H (Eds) Freshwater Invertebrates of the Malaysian Region.Academy of Sciences,Malaysia,191-193pp.
- Use of 18S, 16S rDNA and cytochrome c oxidase sequences in earthworm taxonomy (Oligochaeta, Lumbricidae).Pedobiologia47:428‑433. https://doi.org/10.1016/s0031-4056(04)70217-6
- Pop AA, Csuzdi C, Wink M, Pop VV (2005) An attempt to reconstruct the molecular phylogeny of the genus Allolobophora Eisen, 1874 (sensus lato, Pop, 1941) using 16S rDNA and COI sequences (Oligochaeta, Lumbricidae). In: Pop VV, Pop AA (Eds) Advances in Earthworm Taxonomy II (Annelida: Oligochaeta).Cluj University Press,Cluj-Napoca, Romania,155–164pp.
- Application of 16S, 18S rDNA and COI sequences in the molecular systematics of the earthworm family Lumbricidae (Annelida, Oligochaeta).European Journal of Soil Biology43:S43‑S52. https://doi.org/10.1016/j.ejsobi.2007.08.007
- An annotated checklist of the Romanian earthworm fauna.Zoology in the Middle East58(4):59‑70.
- Analysis of the central nervous system and sense organs in Potamodrilus fluviatilis (Annelida: Potamodrilidae).Zoologischer Anzeiger - A Journal of Comparative Zoology241(1):19‑35. https://doi.org/10.1078/0044-5231-00019
- ZooBank: Developing a nomenclatural tool for unifying 250 years of biological information.Zootaxa1950:39‑50. URL: http://www.mapress.com/zootaxa/2008/f/zt01950p050.pdf
- Contribution à la taxonomie des Hormogastridae (Annelida: Oligochaeta) avec descriptions de nouvelles espèces d'Espagne.Documents pédozoologiques et intégrologiques4:164‑177.
- Elements de paléogéographie lombricienne. Documents pédozoologiques et intégrologiques.Documents pédozoologiques et intégrologiques3:217‑224.
- Révisions des taxons supraspécifiques de Lumbricoidea.Documents pédozoologiques et intégrologiques3:179‑216.
- Sparganophilidae (Annelida, Oligochaeta) distribution in North America and elsewhere, revisited.Megadrilogica12:125‑143.
- Oligochaetes (Clitellata) of the Mata Atlantica (Parana, Brazil): first results of the SOLOBIOMA project.Proceedings of the Estonian Academy of Sciences Biol. Ecol54:302‑309.
- New data on the earthworm fauna of Gallura (northeast Sardinia).Soil Biology and Biochemistry24(12):1383‑1388. https://doi.org/10.1016/0038-0717(92)90122-e
- Rota E (1994a) Enchytraeidae (Annelida: Oligochaeta) of the Mediterranean region: a taxonomic and biogeographic study.National University of Ireland, University College Dublin,255pp.
- Enchytraeidae (Oligochaeta) of western Anatolia: taxonomy and faunistics.Bollettino di Zoologia61(3):241‑260. https://doi.org/10.1080/11250009409355892
- Italian Enchytraeidae (Oligochaeta). I.Boll. Zool62:183‑231. https://doi.org/10.1080/11250009509356067
- Morphology and adaptations of Parergodrilus Reisinger and Hrabeiella Pizl & Chalupský, two enigmatic soil-dwelling annelids.Italian Journal of Zoology65(1):75‑84. https://doi.org/10.1080/11250009809386727
- Oversized enchytraeids (Annelida, Clitellata): a comparative study, with the revised description of Lumbricillus maximus (Michaelsen).Organisms, Diversity and Evolution1:225‑238. https://doi.org/10.1078/1439-6092-00019
- Rota E (2009) Lights on the ground: A historical survey of light production in the Oligochaeta. In: Meyer-Rochow VB (Ed.) Bioluminescence in focus - A collection of illuminating essays.Research Signpost,Trivandrum, Kerala, India,105-138pp.
- Early oligochaete science, from Aristotle to Francesco Redi.Archives of Natural History38(1):136‑163. https://doi.org/10.3366/anh.2011.0011
- From Corsica to Britain: new outdoor records of Ocnerodrilidae (Annelida: Clitellata) in western Europe.Biodiversity Data Journal1(985):1‑12. https://doi.org/10.3897/BDJ.1.e985
- How many lookalikes has Marionina argentea (Michaelsen, 1889) (Annelida: Clitellata: Enchytraeidae)? Three new species described from morphological evidence.Zoologischer Anzeiger252:123‑157. https://doi.org/10.1016/j.jcz.2012.05.001
- Five new species of Enchytraeidae (Annelida: Clitellata) from Mediterranean woodlands of Italy and reaffirmed validity of Achaeta etrusca, Fridericia bulbosa and F. miraflores.Journal of Natural History49:1987‑2020. https://doi.org/10.1080/00222933.2015.1009514
- The enchytraeid fauna of North Africa.Hydrobiologia278:53‑66. https://doi.org/10.1007/978-94-011-0842-3_6
- A taxonomic study of some Swedish Enchytraeidae (Oligochaeta), with descriptions of four new species and notes on the genus Fridericia.Journal of Natural History33(1):29‑64. https://doi.org/10.1080/002229399300461
- Taxonomy and ecology of sponge-associate Marionina spp. (Clitellata: Enchytraeidae) from the Horomatangi geothermal system of Lake Taupo, New Zealand.International Review of Hydrobiology89(1):58‑67. https://doi.org/10.1002/iroh.200310695
- Dichogaster bolaui (Oligochaeta: Octochaetidae), an unusual invader in a swimming pool in Ireland.Journal of Natural History40:161‑167. https://doi.org/10.1080/00222930600630875
- First time in Italy. Is the elusive aquatic megadrile Sparganophilus Benham, 1892 (Annelida, Clitellata) accelerating its dispersal in Europe?Journal of Limnology73(3):482‑489. https://doi.org/10.4081/jlimnol.2014.939
- Community structure, diversity and spatial organization of enchytraeids in Mediterranean urban holm oak stands.European Journal of Soil Biology62:83‑91. https://doi.org/10.1016/j.ejsobi.2014.02.016
- Parergodrilus heideri Reisinger, 1925 (Annelida: Polychaeta) from a holm oak wood in an extinct volcano of southern Italy.Zootaxa2687:65‑68.
- Biogeography and taxonomy of terrestrial Enchytraeidae (Oligochaeta) in northern Sweden, with comparative remarks on the genus Henlea.Zoologischer Anzeiger237(2):155‑169.
- Soil-dwelling polychaetes: enigmatic as ever? Some hints on their phylogenetic relationships as suggested by maximum parsimony analysis of 18S rDNA gene sequences.Contributions to Zoology70:127‑138.
- A molecular phylogeny of annelids.Cladistics23(1):41‑63. https://doi.org/10.1111/j.1096-0031.2006.00128.x
- On some interesting freshwater Annelida, mainly Oligochaeta, of the underground waters of southwestern France with the description of a new species.Subterranean Biology2:1‑5.
- Troglochaetus beranecki Delachaux (Polychaeta, Archiannelida) in esker groundwaters of Finland: a new class of limnic animals for northern Europe.Hydrobiologia379:17‑21. https://doi.org/10.1023/a:1003292202048
- The small annelids (Annelida: Enchytraeidae, Rhyacodrilinae, Aeolosomatidae) in soils of three forests in the White Carpathians (Czech Republic).Acta Soc. Zool. Bohem74(103):115.
- Schlaghamerský J (2014) A brief history of research on potworms (Annelida: Clitellata: Enchytraeidae) of North America. In: Pavlíček T, Cardet P, Almeida MT, Pascoal C, Cássio F (Eds) Advances in Earthworm Taxonomy VI (Annelida: Oligochaeta).Kasparek Verlag,Heidelberg,59-70pp.
- First records of Parergodrilus heideri (Annelida: “Polychaeta”) from North America.Zootaxa3498:81‑86.
- Taxonomy of Fridericia (Oligochaeta, Enchytraeidae). Revision of species with morphological and biochemical methods.Abh. Naturwiss. Ver. Hamburg (N.F.)38:1‑415.
- A guide to European terrestrial and freshwater species of Enchytraeidae (Oligochaeta).Soil Organisms82:1‑176.
- Guide to European terrestrial and freshwater species of Enchytraeidae (Oligochaeta): First supplement.Newsletter on Enchytraeidae12:53‑66.
- Checklist of taxa of Enchytraeidae (Oligochaeta): an update.Soil Organisms87(2):149‑152.
- Diversity of terrestrial Enchytraeidae (Oligochaeta) in Latin America: Current knowledge and future research potential.Applied Soil Ecology69:13‑20. https://doi.org/10.1016/j.apsoil.2012.12.012
- An updated earthworm list for the British Isles and two new ‘exotic’ species to Britain from Kew Gardens.European Journal of Soil Biology45:431‑435. https://doi.org/10.1016/j.ejsobi.2009.07.002
- A classification and the distribution of earthworms, suborder Lumbricina (Haplotaxida: Oligochaeta).Bulletin of the British Museum (Natural History). Zoological Series39:103‑124.
- Earthworms, Synopses of the British Fauna (New Series).Linnean Society of London, The Estuarine and Coastal Sciences Association and Field Studies Council, London31:1‑171.
- The luminous material of Microscolex phosphoreus (Dug.).Biol. Bull.54:191‑195. https://doi.org/10.2307/1537127
- Contributions to the natural history of Lambay, County Dublin.Irish Naturalist16:68‑82.
- Oligochaetes (Naididae, Tubificidae, Opistocystidae, Enchytraeidae, Sparganophilidae and Alluroididae) of Guyana.Hydrobiologia334:17‑29. https://doi.org/10.1007/bf00017350
- Distribution of two species of the earthworm fauna of Šumadija (Serbia) in the Balkans and neighboring territories.Archives of Biological Sciences57(2):133‑136. https://doi.org/10.2298/abs0502133s
- Threat status and distribution of the earthworm genus Helodrilus Hoffmeister, 1845; sensu Zicsi 1985, on the Balkans and the neighboring regions.Biodiversity and Conservation15(14):4601‑4617. https://doi.org/10.1007/s10531-005-5823-9
- Earthworm (Lumbricidae) diversity in the Central Balkans: An evaluation of their conservation status.European Journal of Soil Biology44(1):57‑64. https://doi.org/10.1016/j.ejsobi.2007.09.005
- Earthworms (Oligochaeta: Lumbricidae) of Bulgaria: diversity and biogeographical review.Acta Zoologica Bulgarica2012(4):5‑13.
- Distribution of Lumbricus friendi Cognetti 1904 (Oligochaeta: Lumbricidae) at the European scale: first findings in Serbia.Bulgarian Journal of Agricultural Science20(1):110‑112.
- Diversity and a biogeographical review of the earthworms (Oligochaeta: Lumbricidae) of the Balkan Mountains (Stara Planina Mountains) in Serbia and Bulgaria.Turkish Journal of Zoology37:635‑642. https://doi.org/10.3906/zoo-1301-33
- Ecology and distribution of hyporheic microannelids (Oligochaeta, Aphanoneura , and Polychaeta) from the eastern United States.Arch. Hydrobiol.151(3):493‑510.
- The sister group relationship of Aeolosomatidae and Potamodrilidae (Annelida: “Polychaeta”) — a molecular phylogenetic approach based on 18S rDNA and cytochrome oxidase I.Zoologischer Anzeiger - A Journal of Comparative Zoology243(4):281‑293. https://doi.org/10.1016/j.jcz.2005.01.001
- Detecting possibly saturated positions in 18S and 28S sequences and their influence on phylogenetic reconstruction of Annelida (Lophotrochozoa).Molecular Phylogenetics and Evolution48(2):628‑645. https://doi.org/10.1016/j.ympev.2008.05.015
- The epibiont on crayfishes Hystricosoma chappuisi Michaelsen, 1926 (Oligochaeta, Aeolosomatidae) – new species for Bulgarian fauna.Acta Zoologica Bulgarica32:66‑68.
- Crayfish epibionts Branchiobdella sp. and Hystricosoma chappuisi (Annelida: Clitellata) in Greece.Bull. Fr. Pêche Piscic.387:59‑66. https://doi.org/10.1051/kmae:2007017
- New earthworm records from Bulgaria (Oligochaeta, Lumbricidae).Opuscula Zoologica Budapest44(1):77‑83.
- New earthworm records from the former Yugoslav countries (Oligochaeta, Lumbricidae).Opuscula Zoologica Budapest44(1):61‑76.
- Allolobophora ruzsai sp. n., a new earthworm species and new records from Montenegro (Oligochaeta: Lumbricidae).North Western Journal of Zoology10(1):48‑52.
- New earthworm records from various parts of Greece (Oligochaeta:Lumbricidae, Acanthodrilidae, Megascolecidae, Ocnerodrilidae).Opuscula Zoologica Budapest46(2):143‑152. https://doi.org/10.18348/opzool.2015.2.143
- New earthworm species and records from Albania (Oligochaeta, Lumbricidae).Acta Zoologica Academiae Scientiarum Hungaricae58(3):259‑274.
- New and little known earthworm species from Greece (Oligochaeta: Lumbricidae, Acanthodrilidae).Zootaxa3304:25‑42.
- A new earthworm species and new records from Albania with remarks on the molecular phylogenetic relationships of the Eisenia species of the Balkan Peninsula (Oligochaeta: Lumbricidae).North-Western Journal of Zoology11(1):110‑116.
- New and little known earthworm species from peripheral areas of the Romanian Carpathians (Oligochaeta, Lumbricidae).Acta Zoologica Academiae Scientiarum Hungaricae60(2):85‑107.
- Remarks on the earthworm genus Helodrilus Hoffmeister, 1845 with new epigean and subterranean records (Oligochaeta, Lumbricidae).Opuscula Zoologica Budapest45(2):181‑188.
- Dichogaster bolaui (Michaelsen) (Octochaetidae) - an allochthonous oligochaet invading urban sewer system.Memoranda Societatis pro Fauna et Flora Fennica67:61‑65.
- Propappidae and aquatic Enchytraeidae (Oligochaeta) from the farthest southeast of Russia.Hydrobiologia278:67‑78. https://doi.org/10.1007/bf00142312
- Distribution of freshwater oligochaetes in the west and east coastal regions of the North Pacific Ocean.Hydrobiologia406:67‑81. https://doi.org/10.1007/978-94-011-4207-6_7
- A guide to the freshwater Oligochaeta and Polychaeta of northern and central Europe.Lauterbornia66:1‑235.
- New record of Propappus volki (Annelida: Clitellata: Propappidae) from Japan.Species Diversity11:359‑365.
- Checklist of earthworms (Oligochaeta: Lumbricidae) from Bulgaria—a review.Zootaxa3458:86‑102.
- On the studies of Microdrile Oligochaeta and Aeolosomatidae (Annelida) in China: brief history and species checklist.Acta Hydrobiologica Sinica31:87‑98.
- Evolution of interstitial Polychaeta (Annelida).Hydrobiologia535/536:319‑340. https://doi.org/10.1007/s10750-004-4417-y
- Über eine neue Octodrilus-Art aus Frankreich (Oligochaeta: Lumbricidae).Revue Suisse de Zoologie112:329‑331. https://doi.org/10.5962/bhl.part.80301