|
Biodiversity Data Journal :
Taxonomic Paper
|
|
Corresponding author: Nico Franz (nico.franz@asu.edu)
Academic editor: Li Ren
Received: 12 Sep 2016 | Accepted: 29 Nov 2016 | Published: 03 Feb 2017
© 2017 Nico Franz, Guanyang Zhang
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Franz N, Zhang G (2017) Three new species of entimine weevils in Early Miocene amber from the Dominican Republic (Coleoptera: Curculionidae). Biodiversity Data Journal 5: e10469. https://doi.org/10.3897/BDJ.5.e10469
|
|
Abstract
Background
Using syntactic and semantic conventions of the taxonomic concept approach (
New information
We present detailed images, descriptions, and phylogenetically informed diagnoses for the three new species-level entities, along with logically consistent Region Connection Calculus (RCC-5) alignments of the amended genus-level classifications for Scelianoma Franz and Girón 2009 [FZ2017], Tropirhinus Schoenherr 1823 [FZ2017], and Diaprepes Schoenherr 1823 [FZ2017] - in relation to 2-4 preceding classifications published in 1982-2012. The description of Scelianoma compacta [FZ2017] from Hispaniola is indicative of a more widespread historical range of Scelianoma [FZ2017] than reflected in the extant, southwestern Puerto Rican Scelianoma elydimorpha Franz and Girón 2009 sec.
Keywords
Concept taxonomy, Curculionidae, Dominican amber, Entiminae, fossil, Miocene, new species, weevil
Introduction
The present study adopts the taxonomic concept approach and conventions of
1. Taxonomic concept labels (name sec. author [year];
2. Taxonomic names (without the sec. annotation) are used to refer to any or all usages associated with that name. Example: Diaprepes abbreviatus (Linnaeus 1758).
3. The term [non-focal] is added to taxonomic names whose meanings are not under scrutiny in the present context, such as names for higher-level or outgroup entities. Example: Entiminae Schoenherr, 1823 [non-focal].
For ease of legibility, we abbreviate the often appearing author specifier "sec. Franz & Zhang (2017)" with [FZ2017]. A more expansive justification for these conventions and the associated representations is provided in
We newly name and describe three species-level concepts of broad-nosed weevils (Coleoptera [non-focal]: Curculionidae [non-focal]: Entiminae [non-focal] - higher-level classification in general accordance with
Materials and methods
Provenance of specimen material
The herein utilized specimen material pertains to three amber inclusions on loan from two institutions that serve as their permanent repositories (
1. Snow Entomological Museum Collection, University of Kansas, Lawrence, KS (coden: SEMC; inclusion SEMC 339)
2. Brodzinsky / López-Penha Collection, Department of Paleobiology, National Museum of Natural History, Washington, DC (coden: USNM; inclusions USNM505319 and USNM505325)
These inclusions pertain to the Burdigalian time period of the Early Miocene (Neogene), with an estimated age range of 20.44 ± 0.05 Ma (million years ago) to 15.97 ± 0.05 Ma (
The herein newly designated type specimens have red "holotype" labels that show the genus name and species epithet, gender symbol, author names, year, and source "sec. Franz & Zhang (2017)" (
Imaging and digitization
Imaging. Habitus and detail photographs of the amber-included specimens were produced using either (1) a Leica M205C stereomicroscope, with an attached DFC450 camera, computer, and the Leica Application Suite (LAS) editing software, version 4.1.0.; or (2) a Visionary Digital Passport II sytem and attached Canon EOS Mark 5D II camera. An effort was made to obtain well exposed, fully focused, and scaled images; however the specific conditions of the amber inclusions - including varying optical angles in relation to the amber surface, cracks, and contaminations with non-/organic materials - made this challenging and underscores in importance of assessing the inclusions in person. The images are numbered according to their first appearance in the descriptive section.
Digitization. Darwin Core-compliant information and images for each of the amber-included specimens were added to the "Symbiota Collections of Arthropods Network" (SCAN) portal (see
Systematic analyses
Morphological analysis. Our descriptive approach follows that of preceding treatments of extant West Indian entimine weevils, in particular
Phylogenetic analysis. Inference of the phylogenetic (and hence taxonomic) identity of the specimens was greatly aided by the prior cladistic analysis of
Extension of the character matrix published in Appendix 1 of
| Label / character | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 |
| Scelianoma compacta [FZ2017] | 0 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | – | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Tropirhinus palpebratus [FZ2017] | 0 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| Diaprepes anticus [FZ2017] | 0 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
Extension of the character matrix published in Appendix 1 of
| Label / character | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | 40 | 41 | 42 | 43 | 44 | 45 | 46 | 47 | 48 | 49 | 50 |
| Scelianoma compacta [FZ2017] | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | – | – | 0 | ? | ? |
| Tropirhinus palpebratus [FZ2017] | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | – | – | 0 | ? | ? |
| Diaprepes anticus [FZ2017] | 0 | 0 | 1 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | – | – | 0 | ? | ? |
Extension of the character matrix published in Appendix 1 of
| Label / character | 51 | 52 | 53 | 54 | 55 | 56 | 57 | 58 | 59 | 60 | 61 | 62 | 63 | 64 | 65 | 66 | 67 | 68 | 69 | 70 | 71 | 72 | 73 | 74 | 75 |
| Scelianoma compacta [FZ2017] | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | – | – | 0 | – | 0 |
| Tropirhinus palpebratus [FZ2017] | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | – | – | 0 | – | 0 |
| Diaprepes anticus [FZ2017] | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | – | – | 0 | – | 0 |
Extension of the character matrix published in Appendix 1 of
| Label / character | 76 | 77 | 78 | 79 | 80 | 81 | 82 | 83 | 84 | 85 | 86 | 87 | 88 | 89 | 90 | 91 | 92 | 93 | 94 | 95 | 96 | 97 | 98 | 99 | 100 |
| Scelianoma compacta [FZ2017] | 0 | 0 | 0 | 0 | 0 | – | – | 1 | – | – | 0 | 0 | 0 | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
| Tropirhinus palpebratus [FZ2017] | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ? | ? | ? | 0 | 0 | 0 | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
|
Diaprepes anticus [FZ2017] |
0 | 0 | 0 | 0 | 0 | – | – | 0 | 0 | 0 | 1 | 0 | 0 | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
Extension of the character matrix published in Appendix 1 of
| Label / character | 101 | 102 | 103 | 104 | 105 | 106 | 107 | 108 | 109 | 110 | 111 | 112 | 113 | 114 | 115 | 116 | 117 | 118 | 119 | 120 | 121 | 122 | 123 | 124 | 125 |
| Scelianoma compacta [FZ2017] | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
| Tropirhinus palpebratus [FZ2017] | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
| Diaprepes anticus [FZ2017] | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
Extension of the character matrix published in Appendix 1 of
| Label / character | 126 | 127 | 128 | 129 | 130 | 131 | 132 | 133 | 134 | 135 | 136 | 137 | 138 | 139 | 140 | 141 | 142 | 143 |
| Scelianoma compacta [FZ2017] | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
| Tropirhinus palpebratus [FZ2017] | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
| Diaprepes anticus [FZ2017] | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
In accordance with the preceding analysis (
Taxon treatments
Scelianoma compacta , sp. n.
- SCAN (Symbiota) http://symbiota4.acis.ufl.edu/scan/portal/collections/individual/index.php?occid=25836759
- ZooBank urn:lsid:zoobank.org:act:D123834D-3062-4A6E-81C1-886DD594EF11
-
scientificName: Scelianoma compacta; namePublishedIn:Franz, N.M. & G. Zhang. 2017. Three new species of entimine weevils in Early Miocene amber from the Dominican Republic (Coleoptera: Curculionidae). Biodiversity Data Journal.; nameAccordingTo:Franz & Zhang 2017; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Coleoptera; family:Curculionidae; scientificNameAuthorship:Franz & Zhang, 2017; country:Dominican Republic; stateProvince:La Vega; locality:Unknown mine; decimalLatitude:19; decimalLongitude:-70.666667; geodeticDatum:WGS84; georeferencedBy:N.M. Franz; georeferenceSources:Google Earth; georeferenceVerificationStatus:Verfied by Curator; verbatimEventDate:Early Miocene, Burdigalian; habitat:Amber inclusion, Early Miocene (Burdigalian); individualCount:1; sex:Male; lifeStage:Adult; preparations:Amber inclusion; catalogNumber:ARTSYS0000269; recordNumber:DR-888; recordedBy:Unknown; disposition:SEMC, on loan; otherCatalogNumbers:SEMC339; identifiedBy:N.M. Franz & G. Zhang; dateIdentified:01/15/2017; identificationReferences:Franz, N.M. & G. Zhang. 2017. Three new species of entimine weevils in Early Miocene amber from the Dominican Republic (Coleoptera: Curculionidae). Biodiversity Data Journal.; modified:24/01/2017 18:31; rights: ; rightsHolder:University of Kansas; bibliographicCitation:Franz, N.M. & G. Zhang. 2017. Three new species of entimine weevils in Early Miocene amber from the Dominican Republic (Coleoptera: Curculionidae). Biodiversity Data Journal.; institutionID:KU SEMC; collectionID:0acac5fe-f0ec-4d9f-82f8-0dbb74888de2; institutionCode:SCAN; collectionCode:ARTSYS; ownerInstitutionCode:KU SEMC; basisOfRecord:FossilSpecimen; source: ; occurrenceID:44a28738-bbf7-441d-8343-9adf009eb5b0
Description
Male - habitus (Fig.
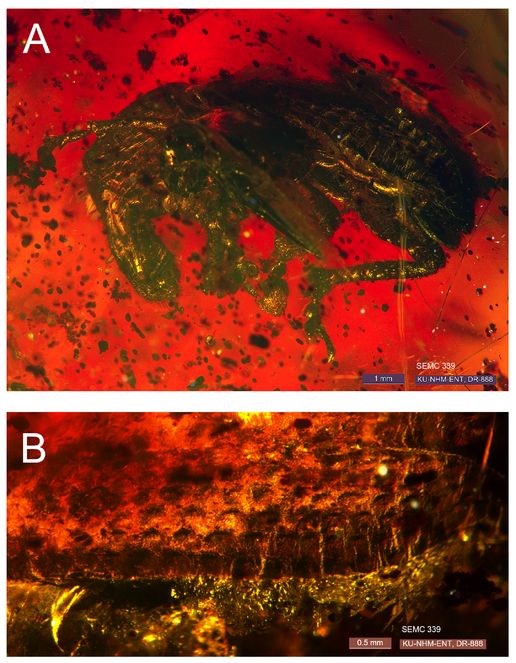
Scelianoma compacta Franz & Zhang 2017 sp. n. [FZ2017], male holotype, specimen SEMC 339 (= ARTSYS0000269). (A) Habitus, left lateral view; (B) Left elytron (incomplete). We note that optical imperfections in the amber inclusion interfere with an entire, undistorted view of the male holotype from a standard viewing angle. Similar imperfections are present in the other inclusions.
Mouthparts. Mandibles equilateral, with 5-8 fine setae; mandibular scar positioned apicolaterally, projected. Maxillae with maxillary palps apparently 3-segmented (2 projecting palpomeres visible). Labium with prementum cordate, slightly wider than long; labial palps apparently 3-segmented.
Rostrum. Length 1.85 mm, rostral/pronotal length ratio 0.6, rostral length/width ratio 2.0 (though rostrum laterally compressed due to fossilization). Rostrum in dorsal view rectangular, anteriorly widened, anterodorsal margin weakly emarginate; epistoma with nasal plate (see
Head. Eyes small, globular (strongly roundly protruded), laterally positioned, separated by distance shorter than anterior-to-posterior length of each eye; outline in lateral profile elliptical (horizontally more extended), anterior and posterior margins more strongly rounded, and with an anteocular invagination (see
Thorax. Pronotum in dorsal view elongate-tubular, length/width ratio 1.5, pronotal/elytral length ratio 0.43; widest near mid point; surface foveate to lacunose (including lateral regions), lacunae somewhat variable in size and arranged in an off-set, honeycomb-like pattern; median sulcus absent. Pronotum in lateral view tubular, slightly arcuate; anterolateral margins straight (without postocular lobe), postocular vibrissae absent. Scutellum exposed by elytra, small, subcircular. Epipleura challenging to observe (distorted, obscured), though apparently similar to those of Scelianoma elydimorpha sec.
Legs. Prothoracic and metathoracic legs each longer than mesothoracic legs, generally similar to those of males of Scelianoma elydimorpha sec.
Elytra (Fig.
Wings. Absent.
Abdomen. Venter with only abdominal ventrites VI and VII visible (and displaced by process of fossilization), each similar in length, and VII with posterior margin widely rounded. Pygidium entirely covered by elytra.
Terminalia. Terminalia not unambiguously observed; however, located just to the left side of the amber-included specimen are several displaced, distorted chitinous structures that apparently include the male spiculum gastrale and median lobe in more or less parallel orientation to the remainder of the specimen. Accordingly (with aforementioned caveats), the presumed spiculum gastrale is similar to that of Scelianoma elydimorpha sec.
Female. Unknown.
Diagnosis
Generic placement. Scelianoma compacta [FZ2017] shares with Scelianoma elydimorpha Franz and Girón sec.
Close extant relatives of Scelianoma Franz and Girón [FZ2017] include members of Artipus Sahlberg sec.
Scelianoma [FZ2017] is also distinct from other extant Caribbean groups such as Apotomoderes Dejean sec.
Differential diagnosis. Scelianoma compacta [FZ2017], in addition to being extinct and recorded from Dominican amber, is differentiated from the extant, southwestern Puerto Rico-inhabiting Scelianoma elydimorpha sec.
Etymology
The epithet - "thick, firm, compact" (
Distribution
Scelianoma compacta [FZ2017] is known only from the examined Dominican amber inclusion ("SEMC 339"; see Material) of the Burdigalian time period. The specific mine of origin for this inclusion is unknown.
Ecology
Unknown.
Tropirhinus palpebratus , sp. n.
- SCAN (Symbiota) http://symbiota4.acis.ufl.edu/scan/portal/collections/individual/index.php?occid=25836760
- ZooBank urn:lsid:zoobank.org:act:2E8D32B1-D021-4E02-BABE-B494098D4C94
-
scientificName: Tropirhinus palpebratus; namePublishedIn:Franz, N.M. & G. Zhang. 2017. Three new species of entimine weevils in Early Miocene amber from the Dominican Republic (Coleoptera: Curculionidae). Biodiversity Data Journal.; nameAccordingTo:Franz & Zhang 2017; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Coleoptera; family:Curculionidae; scientificNameAuthorship:Franz & Zhang, 2017; country:Dominican Republic; stateProvince:La Vega; locality:Unknown mine; decimalLatitude:19; decimalLongitude:-70.666667; geodeticDatum:WGS84; georeferencedBy:N.M. Franz; georeferenceSources:Google Earth; georeferenceVerificationStatus:Verfied by Curator; verbatimEventDate:Early Miocene, Burdigalian; habitat:Amber inclusion, Early Miocene (Burdigalian); individualCount:1; sex:Female; lifeStage:Adult; preparations:Amber inclusion; catalogNumber:ARTSYS0000270; recordNumber:Woodruff #9768; recordedBy:R.E. Woodruff; disposition:USNM, on loan; otherCatalogNumbers:USNM505319; identifiedBy:N.M. Franz & G. Zhang; dateIdentified:01/15/2017; identificationReferences:Franz, N.M. & G. Zhang. 2017. Three new species of entimine weevils in Early Miocene amber from the Dominican Republic (Coleoptera: Curculionidae). Biodiversity Data Journal.; modified:24/01/2017 18:31; rights: ; rightsHolder:United States National Museum; bibliographicCitation:Franz, N.M. & G. Zhang. 2017. Three new species of entimine weevils in Early Miocene amber from the Dominican Republic (Coleoptera: Curculionidae). Biodiversity Data Journal.; institutionID:USNM; collectionID:0acac5fe-f0ec-4d9f-82f8-0dbb74888de2; institutionCode:SCAN; collectionCode:ARTSYS; ownerInstitutionCode:USNM; basisOfRecord:FossilSpecimen; source: ; occurrenceID:266d8782-5bf5-4763-b3fa-ea057a3fc55a
Description
Female - habitus (Fig.

Mouthparts. Mandibles equilateral, with 3-5 coarse and several finer setae; mandibular scar positioned apicolaterally, projected. Maxillae not apparent, covered by labium (however, the maxillary palps are 3-segmented in extant members of Tropirhinus Schoenherr [FZ2017]). Labium with prementum cordate, equilateral; labial palps apparently 3-segmented.
Rostrum. Length 1.3 mm, rostral/pronotal length ratio 0.8, rostral length/width ratio 1.1. Rostrum in dorsal view equilateral to rectangular, dorsolateral margins nearly straight and distance between them anteriorly gradually widening, anterodorsal margin with a distinct, narrow, V-shaped mesal emargination; epistoma with nasal plate (see
Head. Eyes large, globular (strongly roundly protruded), dorsolaterally positioned, separated by distance similar to anterior-to-posterior length of each eye; outline in lateral profile elliptical (horizontally more extended), ventral margin less rounded.
Thorax. Pronotum in dorsal view equilateral to transverse, weakly convex, length/width ratio 0.85, pronotal/elytral length ratio 0.25; widest near posterior 1/3, lateral margins continuously rounded; surface punctate, with a wide, elliptical median sulcus (or impression) extending along anterior 1/2 of pronotum. Pronotum in lateral view equilateral; anterolateral margins with a small postocular lobe, and dorsad thereof with a tuft of 4-6 slightly longer, anteriorly directed setae ("postocular vibrissae", except these are not projecting from the postocular lobe but are dorsad of it). Scutellum exposed by elytra, small, escudate, posterior margins rounded. Epipleura with mespisternum triangular; mesepimeron dorsally oblique truncate; metepisternum linear, anteriorly widened; metepimeron entirely covered by elytron. Prothoracic ventrite with anterior margin widely emarginate; proxocal cavities positioned near mid point, contiguous. Mesothoracic ventrite with plumose-scopiform scales; mesocoxal cavities separeated by distance 1/3 as wide as each mesocoxal cavity. Metathoracic ventrite with median sulcus present as a large, transverse fovea positioned anteriad of posterior margin; metacoxal cavities separated by distance similar to width of each metacoxal cavity. Metendosternite not observed.
Legs. Prothoracic and metathoracic legs each slightly longer than mesothoracic legs (mesofemora shortest in comparison), generally similar to those of Diaprepes abbreviatus sec.
Elytra. Length/width ratio 1.8; widest near mid region; anterior margins jointly wider than posterior margin of pronotum, slightly sinuate; humeri present, rounded; lateral margins continously rounded, nearly straight in mid region, more strongly converging in along posterior 1/4; posterior edges each with a short, narrowly triangular, ante-apical projection. Elytra in lateral view with dorsal outline subplane along anterior 1/2, thereafter continuously rounded (hence declivity convex), less so along posterior 1/8. Elytra with striae I-IX complete, stria X only apparent along anterior and posterior 1/3; striae similar in width to intervals; punctures separated by distance similar to width of each puncture; intervals slightly and roundly elevated; pale-colored scales and setae covering elytra homogenously, with no maculae apparent.
Wings. Present, yet not observed (covered by elytra).
Abdomen. Venter with segments III and IV jointed (see
Terminalia. Not unambiguously observable; however, the stylus and setae of the left coxite appear to project from the terminal opening, which is indicative of the female identity of the specimen (along witht the triangular ventral segment VII).
Male. Unknown.
Diagnosis
Generic placement. Tropirhinus palpebratus [FZ2017] shares with (e.g.) Tropirhinus elegans (Guérin 1847) sec.
We thereby assign to Tropirhinus [FZ2017] an expanded circumscription in comparison to (e.g.) Tropirhinus sec.
Differential diagnosis. Tropirhinus palpebratus [FZ2017], in addition to being extinct and recorded from Dominican amber, is readily distinguished from the extant members of Tropirhinus [FZ2017] by the presence of a small, postocular lobe (with a setal patch ventral thereof) and absence of metallic-colored pronotal and elytral maculae. Moreover, the eyes of Tropirhinus palpebratus [FZ2017] are more globular and protruded than those of Tropirhinus elegans sec.
Etymology
The epithet - "eyelid, wink" (
Distribution
Tropirhinus palpebratus [FZ2017] is known only from the examined Dominican amber inclusion ("USNM505319"; see Material) of the Burdigalian time period. The specific mine of origin for this inclusion is unknown.
Ecology
Unknown.
Diaprepes anticus , sp. n.
- SCAN (Symbiota) http://symbiota4.acis.ufl.edu/scan/portal/collections/individual/index.php?occid=25836761
- ZooBank urn:lsid:zoobank.org:act:92E4FDF2-B9E4-4441-AD74-261EDA89E661
-
scientificName: Diaprepes anticus; namePublishedIn:Franz, N.M. & G. Zhang. 2017. Three new species of entimine weevils in Early Miocene amber from the Dominican Republic (Coleoptera: Curculionidae). Biodiversity Data Journal.; nameAccordingTo:Franz & Zhang 2017; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Coleoptera; family:Curculionidae; scientificNameAuthorship:Franz & Zhang, 2017; country:Dominican Republic; stateProvince:La Vega; locality:Unknown mine; decimalLatitude:19; decimalLongitude:-70.666667; geodeticDatum:WGS84; georeferencedBy:N.M. Franz; georeferenceSources:Google Earth; georeferenceVerificationStatus:Verfied by Curator; verbatimEventDate:Early Miocene, Burdigalian; habitat:Amber inclusion, Early Miocene (Burdigalian); individualCount:1; sex:Female; lifeStage:Adult; preparations:Amber inclusion; catalogNumber:ARTSYS0000271; recordNumber:Woodruff #9774; recordedBy:R.E. Woodruff; disposition:USNM, on loan; otherCatalogNumbers:USNM505325; identifiedBy:N.M. Franz & G. Zhang; dateIdentified:01/15/2017; identificationReferences:Franz, N.M. & G. Zhang. 2017. Three new species of entimine weevils in Early Miocene amber from the Dominican Republic (Coleoptera: Curculionidae). Biodiversity Data Journal.; modified:24/01/2017 18:31; rights: ; rightsHolder:United States National Museum; bibliographicCitation:Franz, N.M. & G. Zhang. 2017. Three new species of entimine weevils in Early Miocene amber from the Dominican Republic (Coleoptera: Curculionidae). Biodiversity Data Journal.; institutionID:USNM; collectionID:0acac5fe-f0ec-4d9f-82f8-0dbb74888de2; institutionCode:SCAN; collectionCode:ARTSYS; ownerInstitutionCode:USNM; basisOfRecord:FossilSpecimen; source: ; occurrenceID:08bb94f8-fddc-4506-b454-34e7d27e5343
Description
Female - habitus (Fig.
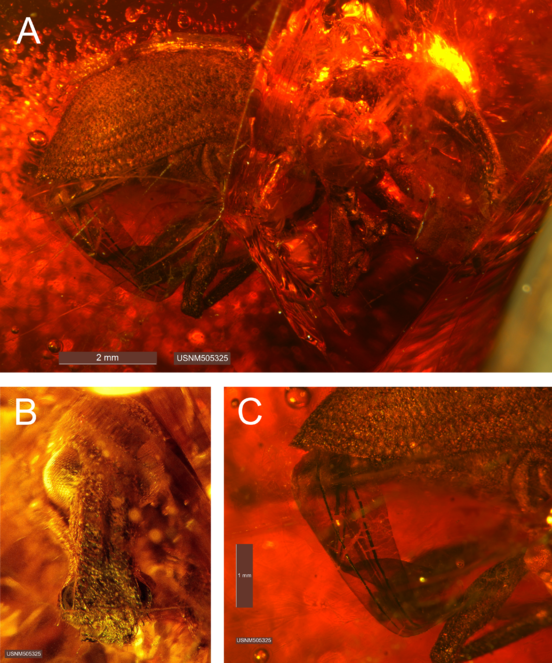
Mouthparts. Mandibles equilateral, asymmetrical, with 6-10 fine setae of variable length; mandibular scar positioned apicolaterally, projected. Maxillae with maxillary palps 3-segmented. Labium with prementum cordate, equilateral; labial palps apparently 3-segmented.
Rostrum. Length 1.45 mm, rostral/pronotal length ratio 0.75, rostral length/width ratio 1.8. Rostrum in dorsal view elongate, dorsolateral margins subparallel and weakly arcuate along posterior 2/3, expanded along anterior 1/3, anterodorsal margin weakly emarginate; epistoma with nasal plate (see
Head. Eyes large, globular (strongly roundly protruded), dorsolaterally positioned, separated by distance slightly shorter than anterior-to-posterior length of each eye; outline in lateral view elliptical (horizontally more extended), ventral margin less rounded.
Thorax. Pronotum in dorsal view equilateral, length/width ratio 1.5 (though challenging to observe due to the fossil's position in an inclusion with limited viewing access of the pronotum), pronotal/elytral length ratio 0.5; widest near posterior margin, lateral margins continously rounded and posteriorly diverging; surface punctate to foveate, with irregularly spaced and shaped concavities, ranging from subcircular to elongate to arcuate (see
Legs. Prothoracic and metathoracic legs each slightly longer than mesothoracic legs (mesofemora shortest in comparison), highly similar to those of Diaprepes abbreviatus sec.
Elytra. Length/width ratio 1.5; widest near mid region; anterior margins jointly wider than posterior margin of pronotum (though challenging to observe along a crack in the amber inclusion), slightly sinuate; humeri present, rounded; lateral margins subparallel along anterior 1/2, therafter gradually and roundly converging, posterior edges narrow, actue, though not projected. Elytra in lateral view with dorsal outline weakly convex along anterior 3/4, thereafter (along posterior 1/4) with weakly angulate, straight declivity. Elytra with striae I-IX complete, stria X only apparent along anterior and posterior 1/3; striae slightly narrower than intervals; punctures separated by distance shorter than or similar to width of each puncture; intervals slightly and roundly elevated, no carinae apparent (as, e.g., in Diaprepes abbreviatus
Wings. Present, and visible in part (apical 1/2) since the specimen had its wings extended prior to its preservation in amber; veins RP1 and RP2 apparent (see
Abdomen. Venter with segments III and IV jointed (see
Terminalia. Not externally visible; however, the triangular, posteriorly narrowly projected ventral segment VII is indicative of this specimen being female (see
Male. Unknown.
Diagnosis
Generic placement. Diaprepes anticus [FZ2017] shares with (e.g.) Diaprepes maugei (Boheman 1840) sec.
Differential diagnosis. Diaprepes anticus [FZ2017], in addition to being extinct and recorded from Dominican amber, is readily distinguished from extant members of Diaprepes [FZ2017] by the absence of postocular vibrissae (character 48[0]), the absence of variously extended, rounded, and glabrate elytra carinae (character 64[0]), and the absence of striped elytral regions with intermixed appressed and suberect scales (character 79[0]; all characters and states according to
Etymology
The epithet - "in front, foremost" (
Distribution
Diaprepes anticus [FZ2017] is known only from the examined Dominican amber inclusion ("USNM505325"; see Material) of the Burdigalian time period. The specific mine of origin for this inclusion is unknown.
Ecology
Unknown.
Discussion
Taxononomic concept alignments
We present consistent Region Connection Calculus (RCC-5) alignments and visualizations of current and preceding taxonomic concepts that are relevant to our newly recognized names and entities. The process of generating such alignments is described in detail in
Alignment of Scelianoma sec. auctorum. (Fig.
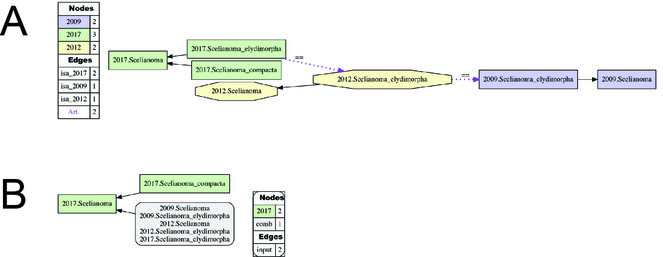
(A) Input visualization and (B) alignment visualization for the taxonomic concept alignment of Scelianoma sec. auctorum. See also Suppl. materials
Alignment of Tropirhinus sec. auctorum. (Fig.
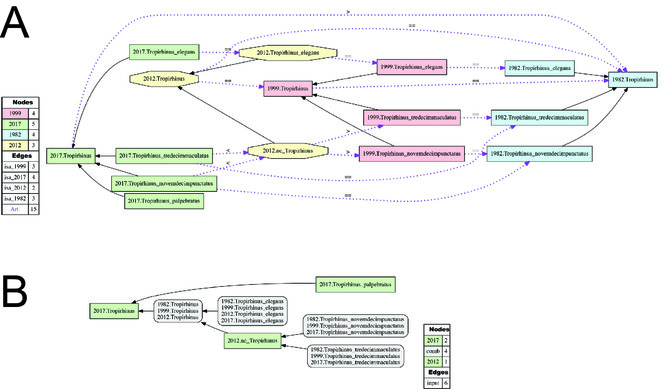
Alignment of Diaprepes sec. auctorum. (Figs
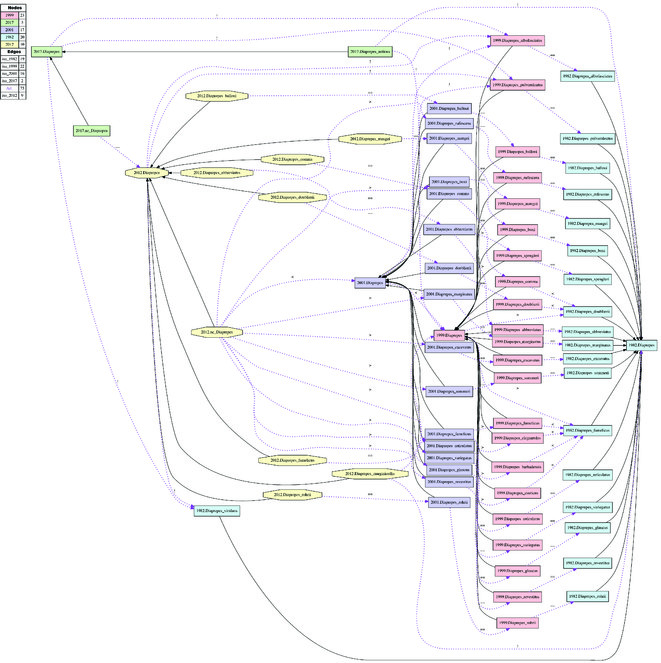
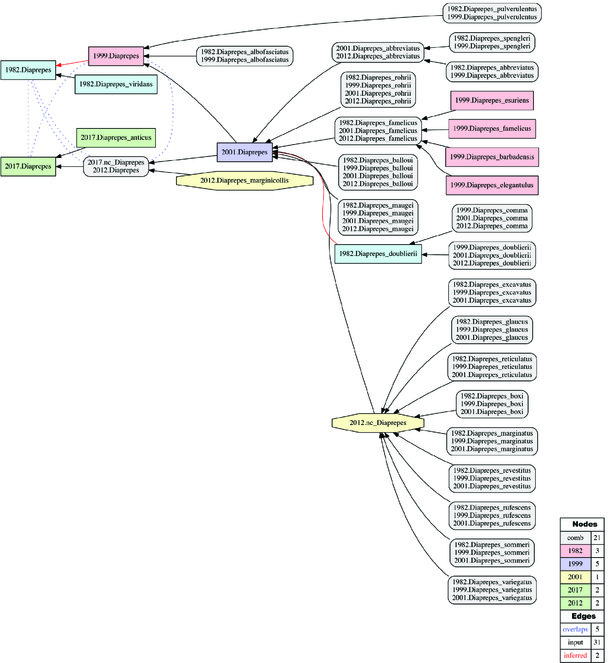
Historical biogeographic implications
The recognition of the three Miocene-based fossils Scelianoma compacta [FZ2017], Tropirhinus palpebratus [FZ2017], and Diaprepes anticus [FZ2017] suggests that the corresponding weevil lineages are longstanding members of a diversified and specialized West Indian weevil fauna (see also
Biodiversity informatics
This study has benefitted from a recent, relevant, and arguably thorough cladistic analysis (
Acknowledgements
The authors thank in particular Steven R. Davis and Michael S. Engel (University of Kansas) for granting access to the herein analyzed fossil specimens (along with Conrad C. Labandeira of the United States National Museum), and also for sharing an earlier, independently generated, and unpublished manuscript in which two of these were described and imaged, although without formal name assignments. This research is supported by the United States National Science Foundation, grants DEB-1155984 and EF-5331207371, and by the United States Department of Agriculture, agreement 58-1275-1-335.
Author contributions
The authors contributed equally to all aspects of this study.
References
- Catalogue of fossil insect species described from Dominican amber (Miocene).Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie)352:1‑68. URL: http://www.naturkundemuseum-bw.de/sites/default/files/publikationen/serie-b/B352.pdf
- The insect and spider collections of the world.Sandhill Crane Press,310pp. [ISBN978-9004081925]
- The concept of "potential taxa" in databases.Taxon44(2):207‑212. URL: http://www.jstor.org/stable/1222443
- Family-Group Names In Coleoptera (Insecta).ZooKeys88:1‑972. https://doi.org/10.3897/zookeys.88.807
- Composition of scientific words, revised edition.Smithsonian Institution Press,Washington, D.C.,882pp.
- Euler/X: a toolkit for logic-based taxonomy integration.arXiv:1402.1992.arXiv. URL: http://arxiv.org/abs/1402.1992
- Descriptions de Coléoptères nouveaux ou peu connus.Annales de la Société Entomologique de France5(7):167‑182. URL: http://biodiversitylibrary.org/page/8244495
- Redescriptions of critical type species in the Eustylini Lacordaire (Coleoptera: Curculionidae: Entiminae).Journal of Natural History44:41‑80. https://doi.org/10.1080/00222930903383495
- Revision and phylogeny of the Caribbean weevil genus Apotomoderes Dejean, 1834 (Coleoptera, Curculionidae, Entiminae).ZooKeys49:33‑75. https://doi.org/10.3897/zookeys.49.303
- Melathra huyenae Franz, a new genus and new species of entimine weevil (Coleoptera: Curculionidae: Entiminae) from southwestern Hispaniola.The Coleopterists Bulletin65(4):352‑362. https://doi.org/10.1649/072.065.0406
- Phylogenetic reassessment of the Exophthalmus genus complex (Curculionidae: Entiminae: Eustylini, Geonemini).Zoological Journal of the Linnean Society164(3):510‑557. https://doi.org/10.1111/j.1096-3642.2011.00774.x
- Anatomy of a cladistic analysis.Cladistics30(3):294‑321. https://doi.org/10.1111/cla.12042
- Description of two new species and phylogenetic reassessment of Perelleschus O'Brien & Wibmer, 1986 (Coleoptera: Curculionidae), with a complete taxonomic concept history of Perelleschus sec. Franz & Cardona-Duque, 2013.Systematics and Biodiversity11(2):209‑236. https://doi.org/10.1080/14772000.2013.806371
- Controlling the taxonomic variable: Taxonomic concept resolution for a southeastern United States herbarium portal.Research Ideas and Outcomes2:e10610. https://doi.org/10.3897/rio.2.e10610
- Reasoning over taxonomic change: exploring alignments for the Perelleschus use case.PLoS ONE10(2):e0118247. https://doi.org/10.1371/journal.pone.0118247
- Names are not good enough: reasoning over taxonomic change in the Andropogon complex.Semantic Web7(6):645‑667. https://doi.org/10.3233/SW-160220
- Two influential primate classifications logically aligned.Systematic Biology65(4):561‑582. https://doi.org/10.1093/sysbio/syw023
- Scelianoma elydimorpha, a new genus and new species of entimine weevil from southwestern Puerto Rico (Coleoptera: Curculionidae, Entiminae).Neotropical Entomology38(2):219‑230. https://doi.org/10.1590/s1519-566x2009000200009
- Phylogenetic assessment of the Caribbean weevil genus Lachnopus Schoenherr (Coleoptera : Curculionidae : Entiminae).Invertebrate Systematics26(1):67. https://doi.org/10.1071/is11033
- TNT, a free program for phylogenetic analysis.Cladistics24(5):774‑786. https://doi.org/10.1111/j.1096-0031.2008.00217.x
- Symbiota – a virtual platform for creating voucher-based biodiversity information communities.Biodiversity Data Journal2:e1114. https://doi.org/10.3897/bdj.2.e1114
- Evolution of the Insects.1.Cambridge University Press,xv + 755pp. [ISBN978-0521821490]
- Décade entomologique.Revue Zoologique10:2‑11. URL: http://biodiversitylibrary.org/page/2271611
- Hilgen FJ, Lourens LJ, Dam JAV, Beu AG, Boyes AF, Cooper RA, Krijgsman W, Ogg JG, Piller WE, Wilson DS (2012) The Neogene Period. The Geologic Time Scale.Elsevier,923-978pp. https://doi.org/10.1016/b978-0-444-59425-9.00029-9
- Phylogenetic revision of Minyomerus sec. Jansen & Franz, 2015 (Coleoptera, Curculionidae) using taxonomic concept annotations and alignments.ZooKeys528:1‑133. https://doi.org/10.3897/zookeys.528.6001
- The ventral structures of the weevil head (Coleoptera: Curculionoidea).Memoirs of the Entomological Society of Washington14:35‑51.
- The species of Entiminae (Coleoptera: Curculionidae) ranged in America south of the United States.Anales del Instituto de Biología, Universidad Nacional Autónoma de México. Serie Zoología70(2):99‑168. URL: http://www.redalyc.org/articulo.oa?id=45870204
- ASADO, version 1.85 TNT-mrbayes Slaver version 2; mxram 200 (vol 5.30).Made available through the author, Cornell University, Ithaca, NY.
- Oberprieler R, Anderson R, Marvaldi A (2014) Curculionoidea Latreille, 1802: introduction, phylogeny; pp. 285-300. In: Leschen RB, Beutel R (Eds) Handbook of Zoology: Arthropoda: Insecta: Coleoptera: Volume 3: Morphology and Systematics (Phytophaga).de Gruyter,Berlin,XI + 673pp. [ISBN978-3-11-027446-2].
- Diaprepes, Short Course & Workshop Proceedings, Edited by Stephen H. Futch.Citrus Research and Education Center, University of Florida Institute of Food and Agricultural Sciences, Lake Alfred.1-7pp. URL: http://irrec.ifas.ufl.edu/flcitrus/short_course_and_workshop/diaprepes/genus_diaprepes.shtml
- Annotated checklist of the weevils (Curculionidae sensu lato) of North America, Central America, and the West Indies (Coleoptera: Curculionoidea).Memoirs of the American Entomological Institute34:1‑382.
- Palaeoecological perspectives in Dominican amber.Annales de la Société entomologique de France (N.S.)46:23‑52. https://doi.org/10.1080/00379271.2010.10697637
- First record of the genus Baris Germar, 1817 (Coleoptera: Curculionidae), in Dominican amber.Fossil Record18(1):31‑35. https://doi.org/10.5194/fr-18-31-2015
- Brachycamacina, a new subtribe of the tribe Naupactini (Coleoptera: Curculionidae Entiminae) in Dominican amber.Palaeontologia Electronica16:3:1‑9. URL: http://palaeo-electronica.org/content/2013/531-new-weevil-tribe
- The West Indies as a laboratory of biogeography and evolution.Philosophical Transactions of the Royal Society B: Biological Sciences363(1502):2393‑2413. https://doi.org/10.1098/rstb.2007.2068
- Entrapment bias of arthropods in Miocene amber revealed by trapping experiments in a tropical forest in Chiapas, Mexico.PLOS ONE10:3:e0118820. https://doi.org/10.1371/journal.pone.0118820
- Observations on the morphology and classification of weevils (Coleoptera, Curculionoidea) with a key to major groups.Journal of Natural History26(4):835‑891. https://doi.org/10.1080/00222939200770511
- A revision of the South American genus Hyphantus (Coleoptera, Curculionidae, Otiorhynchinae).Bulletin of the American Museum of Natural History125(4):241‑304. URL: http://hdl.handle.net/2246/1979
- To name or not to name: criteria to promote economy of change in Linnaean classification schemes.Zootaxa3636(2):201‑244. https://doi.org/10.11646/zootaxa.3636.2.1
- Darwin Core: an evolving community-developed biodiversity data standard.PLoS ONE7(1):e29715. https://doi.org/10.1371/journal.pone.0029715
- New genus and species of broad-nosed weevils from Baltic amber and notes on fossils of the subfamily Entiminae (Coleoptera, Curculionidae).ZooKeys160:73‑96. https://doi.org/10.3897/zookeys.160.2108
- Model selection in statistical historical biogeography of Neotropical insects - the Exophthalmus genus complex (Curculionidae: Entiminae).Molecular Phylogenetics and Evolution109:226‑239. https://doi.org/10.1016/j.ympev.2016.12.039
Supplementary materials
Euler/X input data file for the alignment of Scelianoma sec. 2009, 2012, and 2017.
Euler/X output of 16 Maximally Informative Relations inferred for the Scelianoma alignment
Euler/X input data file for the alignment of Tropirhinus sec. 1982, 1999, 2012, and 2017.
Euler/X output of 95 Maximally Informative Relations inferred for the Tropirhinus alignment
Euler/X input data file for the alignment of Diaprepes sec. 1982, 1999, 2001, 2012, and 2017.
Euler/X output of 2001 Maximally Informative Relations inferred for the Diaprepes alignment