|
Biodiversity Data Journal :
Research Article
|
|
Corresponding author: Danwei Huang (huangdanwei@nus.edu.sg)
Academic editor: Pavel Stoev
Received: 05 Dec 2016 | Accepted: 07 Feb 2017 | Published: 13 Feb 2017
© 2017 Rosa Poquita-Du, Chin Soon Lionel Ng, Jun Bin Loo, Lutfi Afiq-Rosli, Ywee Chieh Tay, Peter Todd, Loke Ming Chou, Danwei Huang
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Poquita-Du R, Ng C, Loo J, Afiq-Rosli L, Tay Y, Todd P, Chou L, Huang D (2017) New evidence shows that Pocillopora‘damicornis-like’ corals in Singapore are actually Pocillopora acuta (Scleractinia: Pocilloporidae). Biodiversity Data Journal 5: e11407. https://doi.org/10.3897/BDJ.5.e11407
|
|
Abstract
Molecular sequence data have previously revealed the existence of cryptic species associated with the Pocillopora ‘damicornis-like’ coral. Recently, this species complex has been reclassified into three species including the resurrected P. acuta, which appears to have a wide distribution. Morphological characteristics described for P. acuta are present in corals previously identified as Pocillopora damicornis. To determine if the Pocillopora ‘damicornis-like’ colonies on Singapore’s reefs are P. acuta, P. damicornis, or both, we examined a new collection of Pocillopora using mitochondrial DNA data and morphology. We also compared specimen morphologies from past collections and examined the known regional distributions of both species. Our analyses show that most Pocillopora ‘damicornis-like’ corals in Singapore are P. acuta instead of P. damicornis. Findings here are important for coral diversity records in Singapore and will help clarify distributional limits of morphologically similar Pocillopora species.
Keywords
biodiversity, cryptic species, distribution, museum specimens, phylogenetic analysis, Pocillopora
Introduction
Numerous hard coral species exhibit substantial intraspecific morphological variation (
The genus Pocillopora Lamarck, 1816, is of special interest as it is characterised by exceptionally high levels of phenotypic variation (
Recently, P. acuta was re-established as an entirely separate species from P. damicornis based on differences in their mitochondrial open reading frame (ORF) sequences (
The morphological characteristics described for P. acuta are exhibited by corals previously identified as Pocillopora damicornis in Singapore. Based on this observation, we hypothesise that most of the Pocillopora colonies on Singapore’s reefs are likely to be P. acuta and not P. damicornis. Here, we examine a range of P. ‘damicornis-like’ (sensu
Results of this study are important for coral diversity records in Singapore and will also help clarify the geographical range limits of morphologically closely-related Pocillopora species. As one of the most widespread corals on Singapore's reefs (
Materials and methods
Sixteen Pocillopora samples were collected from five coral reef sites across the southern offshore islands of Singapore. We focused on a wide range of colony morphologies that, following

Pocillopora specimens examined in this study. In situ appearances (A: HD159, D: HD162, G: HD161, J: HD160, M: HD154), with corresponding images of bleached skeletons (B, E, H, K, N). C, live specimen showing brown ring surrounding each oral opening (image by Jenny). F, I, branches from colonies shown in D and G respectively. L, O, calices and septa from colonies shown in J and M respectively. Scale bars represent 1 cm (B, E, H, K, N) and 1 mm (F, I, L, O) respectively.

Pocillopora specimens previously identified as P. damicornis from the Zoological Reference Collection, Lee Kong Chian Natural History Museum, Singapore (A, B: ZRC.1980.20.133; C, D: ZRC.1991.766; E, F: ZRC.1987.1538; G, H: ZRC.1987.1995; I, J: ZRC.1991.763; K, L: ZRC.1987.1537). A–F, colonies with thick branches; G–L, colonies with thinner branches. Scale bars represent 1 cm.
DNA was extracted by overnight digestion in hexadecyltrimethylammonium bromide (CTAB) and proteinase K, followed by phase separation using phenol: chloroform: isoamyl-alcohol (25:24:1). Polymerase chain reactions primed using FATP6.1 and RORF were performed according to
We compiled in Mesquite 3.10 (
We used RAxML 8.0.9 (
Results
Our phylogenetic analysis of seven Pocillopora species recovers two moderately-supported monophyletic groups of P. meandrina + P. verrucosa (P. damicornis type γ) and ‘Clade 1’, as defined by
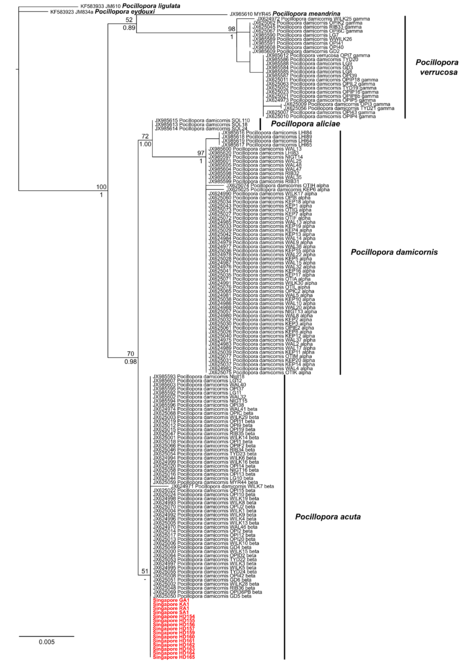
The topology and statistical supports within this clade match those obtained by
Pocillopora acuta Lamarck, 1816
Pocillopora acuta Lamarck, 1816, p. 274;
Materials examined. MNHN-IK-2010-792 (holotype, Muséum national d’Histoire naturelle de Paris, France; type locality: Indian Ocean); see Table
|
Specimen no. |
Catalogue no. |
Locality |
Latitude, Longitude |
Date collected |
Collector |
Last identification |
|
RA1 |
- |
Raffles Lighthouse |
|
Oct 2015 |
R.C. Poquita-Du |
Pocillopora damicornis |
|
GA1 |
- |
Raffles Lighthouse |
|
Oct 2015 |
R.C. Poquita-Du |
Pocillopora damicornis |
|
SA1 |
- |
St. John’s Island |
|
Oct 2015 |
R.C. Poquita-Du |
Pocillopora damicornis |
|
HD154 |
ZRC.CNI.1067 |
Pulau Subar Darat |
|
Oct 2016 |
D. Huang |
Pocillopora acuta |
|
HD155 |
- |
Pulau Subar Darat |
|
Oct 2016 |
D. Huang |
Pocillopora acuta |
|
HD156 |
- |
Pulau Subar Darat |
|
Oct 2016 |
Y.C. Tay |
Pocillopora acuta |
|
HD157 |
ZRC.CNI.1068 |
Pulau Subar Darat |
|
Oct 2016 |
Y.C. Tay |
Pocillopora acuta |
|
HD158 |
- |
Pulau Subar Darat |
|
Oct 2016 |
Y.C. Tay |
Pocillopora acuta |
|
KA1 |
- |
Kusu Island |
|
Oct 2015 |
R.C. Poquita-Du |
Pocillopora damicornis |
|
HD159 |
ZRC.CNI.1069 |
Kusu Island |
|
Oct 2016 |
C.S.L. Ng |
Pocillopora acuta |
|
HD160 |
ZRC.CNI.1070 |
Kusu Island |
|
Oct 2016 |
C.S.L. Ng |
Pocillopora acuta |
|
HD161 |
ZRC.CNI.1071 |
Pulau Subar Laut |
|
Oct 2016 |
Y.C. Tay |
Pocillopora acuta |
|
HD162 |
ZRC.CNI.1072 |
Pulau Subar Laut |
|
Oct 2016 |
Y.C. Tay |
Pocillopora acuta |
|
HD163 |
- |
Pulau Subar Laut |
|
Oct 2016 |
Y.C. Tay |
Pocillopora acuta |
|
HD164 |
- |
Pulau Subar Laut |
|
Oct 2016 |
Y.C. Tay |
Pocillopora acuta |
|
HD165 |
- |
Pulau Subar Laut |
|
Oct 2016 |
Y.C. Tay |
Pocillopora acuta |
|
- |
ZRC.1980.3.20.133 |
Sentosa |
- |
Sep 1979 |
L.T. Chan |
Pocillopora damicornis |
|
- |
ZRC.1987.1537 |
Pulau Hantu |
- |
1987 |
L.M. Chou |
Pocillopora damicornis |
|
- |
ZRC.1987.1538 |
Pulau Hantu |
- |
1987 |
L.M. Chou |
Pocillopora damicornis |
|
- |
ZRC.1991.763 |
Pulau Hantu |
- |
1991 |
Reef Ecology Study Team |
Pocillopora damicornis |
|
- |
ZRC.1991.766 |
Singapore |
1991 |
Reef Ecology Study Team |
Pocillopora damicornis |
|
|
- |
ZRC.1987.1995 |
Singapore |
- |
- |
Reef Ecology Study Team |
Pocillopora damicornis |
Description. Colonial, densely caespitose (Fig.
Remarks. Colonies collected from Singapore’s reefs show great variation in branching morphologies, overlapping with those described for Pocillopora acuta and P. damicornis by
Discussion
This study contributes to the limited data that have emerged from the South China Sea region on the identity of Pocillopora species. The recently-revived species P. acuta is described to have a wide distribution reaching from the central Pacific to the Indian Ocean (
Although we find no contemporary evidence of P. damicornis on Singapore’s reefs, the variability of colony branch thickness among specimens from past collections held at the LKCNHM (Fig.
The taxonomy of South China Sea Pocillopora remains poorly understood. A previous study has shown that Pocillopora ‘damicornis-like’ types 4 and 5 are present in Taiwan, while the Gulf of Thailand only hosts the latter type (
Broadly, Pocillopora acuta is present in both Taiwan and Gulf of Thailand, but P. damicornis appears to be limited to the northern South China Sea as it has thus far only been confirmed from Taiwan using the mitochondrial ORF. Further north at the Yaeyama Islands, Japan, P. damicornis is present but is likely rare relative to P. acuta (
Thus far ranging from the Central Pacific to the Indian Ocean through Singapore, Taiwan and Hawai’i, further sampling may reveal the presence of P. acuta in more localities in the Central Indo-Pacific. Overall, the emerging picture shows that most ‘damicornis-like’ corals in the southwestern South China Sea region are actually P. acuta instead of P. damicornis.
Acknowledgements
This study is funded by the National Research Foundation (Singapore) through the Marine Science R&D Programme (R-154-000-A25-281). We thank Bert Hoeksema and an anonymous reviewer for helping to improve the manuscript, and Chua Keng Soon (LKCNHM) for assistance with loans, imaging and cataloguing of specimens.
References
- Species concept and ecomorph variation in living and fossil Scleractinia.Palaeontographica Americana54:70‑79. URL: http://archive.org/details/palaeontographic541984pale
- Chou LM (2006) Marine habitats in one of the world’s busiest harbours. In: Wolanski E (Ed.) The Environment in Asia Pacific Harbours.Springer,The Netherlands,377-391pp. https://doi.org/10.1007/1-4020-3655-8_22
- Planulation in the scleractinian coral Pocillopora damicornis in Singapore waters.Proceedings of the 7th International Coral Reef Symposium1:500.
- jModelTest 2: more models, new heuristics and parallel computing.Nature Methods9(8). https://doi.org/10.1038/nmeth.2109
- Molecular phylogeny and systematics of the scleractinian coral genus Pocillopora in Hawaii.Proceedings of the 10th International Coral Reef Symposium24‑29.
- The mitochondrial genome of Pocillopora (Cnidaria: Scleractinia) contains two variable regions: The putative D-loop and a novel ORF of unknown function.Gene401:80‑87. https://doi.org/10.1016/j.gene.2007.07.006
- Patterns of genetic structure among Hawaiian corals of the genus Pocillopora yield clusters of individuals that are compatible with morphology.Comptes Rendus Biologies331:239‑247. https://doi.org/10.1016/j.crvi.2007.12.003
- Patterns of small-scale variation of skeletal morphology within the scleractinian corals, Montastrea annularis and Siderastrea siderea.Proceedings of the Third International Coral Reef Symposium2:409‑415. URL: http://www.reefbase.org/resource_center/publication/pub_22034.aspx
- The relationship between corallite morphology and colony shape in some massive reef-corals.Coral Reefs2:19‑25. https://doi.org/10.1007/BF00304728
- A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood.Systematic Biology52(5):696‑704. https://doi.org/10.1080/10635150390235520
- An inventory of zooxanthellate scleractinian corals in Singapore, including 33 new records.Raffles Bulletin of ZoologySupplement 22:69‑80. URL: http://rmbr.nus.edu.sg/rbz/biblio/s22/s22rbz069-080.pdf
- MRBAYES: Bayesian inference of phylogenetic trees.Bioinformatics17(8):754‑755. https://doi.org/10.1093/bioinformatics/17.8.754
- MAFFT multiple sequence alignment software version 7: improvements in performance and usability.Molecular Biology and Evolution30(4):772‑780. https://doi.org/10.1093/molbev/mst010
- Recent developments in the MAFFT multiple sequence alignment program.Briefings in Bioinformatics9(4):286‑298. https://doi.org/10.1093/bib/bbn013
- MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform.Nucleic Acids Research30(14):3059‑3066. https://doi.org/10.1093/nar/gkf436
- Correlated evolution of sex and reproductive mode in corals (Anthozoa: Scleractinia).Proceedings of the Royal Society B-Biological Sciences278(1702):75‑81. https://doi.org/10.1098/rspb.2010.1196
- Most Pocillopora damicornis around Yaeyama Islands are Pocillopora acuta according to mitochondrial ORF sequences.Galaxea17(1):21‑22. https://doi.org/10.3755/galaxea.17.21
- Trapezia septata Dana, 1852 (Brachyura, Trapeziidae): a new record for Singapore with notes on its relationship with the host coral, Pocillopora verrucosa.Crustaceana82(12):1603‑1608. https://doi.org/10.1163/156854009X463838
- The role of taxonomy in species conservation.Philosophical Transactions of the Royal Society B: Biological Sciences359(1444):711‑719. https://doi.org/10.1098/rstb.2003.1454
- Mesquite: A Modular System for Evolutionary Analysis (Version 3.10). URL: http://mesquiteproject.org
- Extreme phenotypic polymorphism in the coral genus Pocillopora; micro-morphology corresponds to mitochondrial groups, while colony morphology does not.Bulletin of Marine Science90(1):211‑231. https://doi.org/10.5343/bms.2012.1080
- A survey of pocilloporid corals and their endosymbiotic dinoflagellate communities in the Austral and Cook Islands of the South Pacific.Platax12:1‑17.
- Coral restoration in Singapore’s sediment-challenged sea.Regional Studies in Marine Science8:422‑429. https://doi.org/10.1016/j.rsma.2016.05.005
- Blind to morphology: genetics identifies several widespread ecologically common species and few endemics among Indo-Pacific cauliflower corals (Pocillopora, Scleractinia).Journal of Biogeography40:1595‑1608. https://doi.org/10.1111/jbi.12110
- jModelTest: Phylogenetic model averaging.Molecular Biology and Evolution25(7):1253‑1256. https://doi.org/10.1093/molbev/msn083
- Tracer: MCMC Trace Analysis Tool. Version 1.6. URL: http://beast.bio.ed.ac.uk/Tracer
- MrBayes 3: Bayesian phylogenetic inference under mixed models.Bioinformatics19(12):1572‑1574. https://doi.org/10.1093/bioinformatics/btg180
- MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space.Systematic Biology61(3):539‑542. https://doi.org/10.1093/sysbio/sys029
- With eyes wide open: a revision of species within and closely related to the Pocillopora damicornis species complex (Scleractinia; Pocilloporidae) using morphology and genetics.Zoological Journal of the Linnean Society170(1):1‑33. https://doi.org/10.1111/zoj.12092
- Daytime spawning of Pocillopora species in Kaneohe Bay, Hawai'i.Galaxea16:11‑12. https://doi.org/10.3755/galaxea.16.11
- Broadcast spawning by Pocillopora species on the Great Barrier Reef.PLoS ONE7(12). https://doi.org/10.1371/journal.pone.0050847
- Assessing hidden species diversity in the coral Pocillopora damicornis from Eastern Australia.Coral Reefs32(1):161‑172. https://doi.org/10.1007/s00338-012-0959-z
- RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models.Bioinformatics22(21):2688‑2690. https://doi.org/10.1093/bioinformatics/btl446
- RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies.Bioinformatics30(9):1312‑1313. https://doi.org/10.1093/bioinformatics/btu033
- A rapid bootstrap algorithm for the RAxML web servers.Systematic Biology57(5):758‑771. https://doi.org/10.1080/10635150802429642
- Genotype × environment interactions in transplanted clones of the massive corals Favia speciosa and Diploastrea heliopora.Marine Ecology-Progress Series271:167‑182. https://doi.org/10.3354/meps271167
- Spatial variability of epibiotic assemblages on marina pontoons in Singapore.Urban Ecosystems20:183‑197. https://doi.org/10.1007/s11252-016-0589-2
- Heterotrophy in recruits of the scleractinian coral Pocillopora damicornis.Marine and Freshwater Behaviour and Physiology46(5):313‑320. https://doi.org/10.1080/10236244.2013.832890
- Corals in Space and Time.UNSW Press,Sydney,320pp.
- Corals of the World.Australian Institute of Marine Science,Townsville,1381pp.
- Scleractinia of Eastern Australia. Part I. Families Thamnasteriidae, Astrocoeniidae, Pocilloporidae.1.Australian Institute of Marine Science,Townsville,85pp. URL: http://archive.org/details/ScleractiniaEas00JENVB
- Taxonomic triage and the poverty of phylogeny.Philosophical Transactions of the Royal Society B: Biological Sciences359(1444):571‑583. https://doi.org/10.1098/rstb.2003.1452