|
Biodiversity Data Journal :
Research Article
|
|
Corresponding author: Evelyne T Lennette (elennette@nectandra.org)
Academic editor: Charles Zartman
Received: 12 Jan 2017 | Accepted: 20 Mar 2017 | Published: 24 Mar 2017
© 2017 Daniel Norris, Ekaphan Kraichak, Allen Risk, Diane Lucas, Dorothy Allard, Frida Rosengren, Theresa Clark, Nicole Fenton, Michael Tessler, Nonkululo Phephu, Evelyne Lennette
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Norris D, Kraichak E, Risk A, Lucas D, Allard D, Rosengren F, Clark T, Fenton N, Tessler M, Phephu N, Lennette E (2017) On the diversity and richness of understory bryophytes at Nectandra Cloud Forest Reserve, Costa Rica. Biodiversity Data Journal 5: e11778. https://doi.org/10.3897/BDJ.5.e11778
|
|
Abstract
Background
A survey of the understory bryophytes in the Nectandra Cloud Forest Preserve yielded 1083 specimens distributed among 55 families, represented by 74 genera of mosses, 75 genera of liverworts and 3 of hornworts. We studied and analyzed the bryophytic distribution on six types of substrates: 1) corticolous, 2) epiphyllous, 3) saxicolous, 4) terricolous, 5) aquatic and 6) lignicolous. The richness and composition of bryophyte genera are compared to those of other previous bryophyte surveys from 4 other sites with different oceanic exposures, climatic and geographic conditions in Costa Rica.
New information
This is a report of the first extensive general survey of bryophytes at the Nectandra Reserve, a premontane cloud forest located on the Atlantic slope of Costa Rica, an area much less studied compared to the Monteverde cloud forest on the Pacific slope.
Keywords
Tropical bryophytes, bryophyte surveys, mosses, liverworts, Nectandra Cloud Forest Reserve, premontane cloud forest, Costa Rica
Introduction
Costa Rica’s climate and weather are determined by the Atlantic and Pacific oceanic influences driven across a very narrow landmass with a backbone of volcanic ranges. Orographic uplifting of the dominant northeasterly Atlantic wet trade winds, which directly impinge against the Cordillera Tilarán, result in intense precipitation on the mid-to-upper Atlantic slope (1000-3000m) in the form of unremitting rain and wind-driven cloud during most of the year, peaking in December-February. In comparison, the west-facing Pacific slopes are steeper, on the leeward side of the trade winds, hence are drier with fractured regions of cloud coverage (
The richness of tropical bryophytes in cloud forests is overwhelming. Bryophytes are found mostly in complex tangles, in long, heavy aerial strands, or in thick mats on all surfaces starting from the ground all the way to the top of the forest canopy, with an abundance of epiphyllous bryophytes in between. As a result, bryophyte ecology is not well studied and its enormous diversity is only beginning to be appreciated. Increasing number of recent reports highlighted the multiple and complex ecologic roles the bryophytes play in the tropical forest. Through poikilohydry, bryophytes resist desiccation and trap water after rehydration. They retain, fix and cycle free atmospheric inorganic nitrogen, carbon and ions (
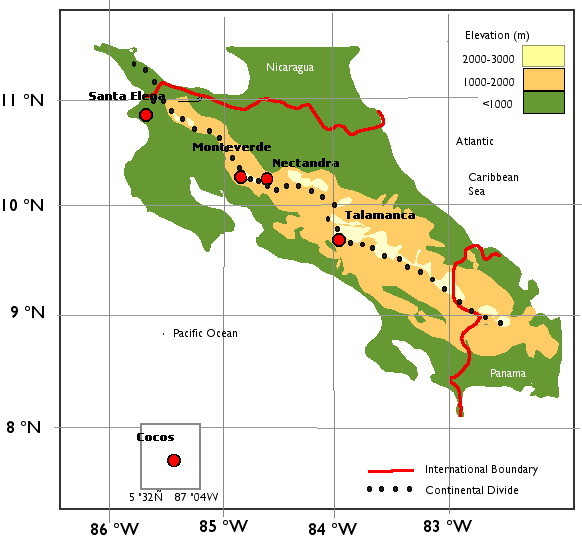
Current area-based floristic information of Costa Rican bryophytes can be gleaned from published, general surveys at four main localities (Fig.
Detailed botanic studies at the Monteverde Cloud Forest Reserve showed that the vegetation richness on the Pacific slope is slightly over half of that on the wetter Atlantic slope, where plant diversity increases with the moisture gradient from mid-to-high (700-1500 m) elevation (
Nectandra Cloud Forest Reserve (henceforth Nectandra) is a private reserve dedicated to cloud forest conservation on the Atlantic slope of the Cordillera de Tílarán, at 1100 -1200m elevation. It is located 40km southeast of the Monteverde Cloud Forest Reserve. Nectandra’s Atlantic exposure, lower elevation, and proximity to Monteverde Cloud forest Reserve presented us with the opportunity to compare the bryoflora of the lower but wetter Atlantic slopes to that of the more studied Monteverde Cloud Forest Reserve. At Nectandra, the higher average precipitation, more moderate temperature and lower elevation are all favorable conditions for higher bryodiversity compared to the Monteverde Reserve. Our survey will hopefully provide a useful comparison of bryoflora from five databases for future research on the effects of climate change on cloud forest.
Materials and Methods
Study Area
The 158 ha Nectandra Reserve (
Nectandra is subclassified as tall-statured montane cloud forest based on biologic and hydrometeorologic variables (
Vascular Plant Communities
The vascular plant diversity was determined from 30 permanent survey plots (10m x 20m) randomly distributed over the entire property (unpublished data). All the vascular plants in each plot were identified, tallied and tagged for monitoring. Of the total of 918 trees, at least 128 distinct species were identified. The most dominant plants in order of decreasing abundance included tree ferns Alsophila firma, Cyathea schiediana, Alsophila imrayana), non-ferns Guettarda poasana, Ocotea tonduzii, Conostegia oerstediana, Elaegia auriculata, each with species density averaging at least two individuals per plot. The 26 genera with the largest trees include Guarea (range 47–104 cm in diameter), Dussia, Hedyosmum (44-100 cm), Ficus (47-80 cm), Ocotea (42-76 cm) , Guatteria (60-75 cm), Paquira (40-73 cm), and Billia (53 – 76 cm) .
Sampling Method
Between 2007 and 2009, D.N. made two separate surveys of mainly understory bryophytes at Nectandra. A third effort (2010) was made in conjunction with a tropical bryophyte course, taught by D.N and E.K. It was attended by eleven participants in the Bryophyte Study Group from seven countries, eight of whom contributed to the collection.
Haphazard floristic sampling of mosses, liverworts and lichens was carried out within 4m of 10 km of trails (equivalent sampled area of 4 ha) on all microhabitats ≤2m in height, including soil, soil banks, streams, rocks, tree trunks, branches, twigs, living leaves. The bryophyte growth on all surfaces is typically dense and entangled. One to five gram-size clumps or mats of candidate specimens were removed from the hard or pendant surfaces (bark/trunk, rock/asphalt) and placed in pre-numbered packets for evaluation and identification. The morphology of bryophyte species were examined in the laboratory with Zeiss dissecting and compound microscopes. With mixed clumps, the species of interest was teased away from the main clump and placed in an individual micro-packet for identification. Multiple micro-packets containing different, segregated, individual species of interest were prepared for each clump/leaf collected. Each outer packet hence contained the remainder of the clump and the associated micro-packets of the voucher specimens for the herbarium. The micro-packets in each main packet were differentiated by letters of the alphabet. The collection (see Supplementary material list A) was deposited at the Jepson Herbarium at the University of California at University of California, Berkeley. At the time of this report, 75% of the collection have been accessioned and the sample information are accessible online through www.ucjepson. Only bryophyte specimens with sufficient information and identification, completed to the level of genera were included in this report. To evaluate the completeness of the collection, we calculated the projected number of species, using the species accumulation curve (the function “specaccum”) and the species pool richness estimator (the function “specpool”) in the R-package “vegan”, from the accessions that were identified to the species level (
The identification of the specimens followed the key and description provided in
Results
General Characteristics of Samples
There were a total of 1083 specimens examined at the genus level (Suppl. material
Only a subset of 488 specimens were identified with confidence to the species level (Suppl. material
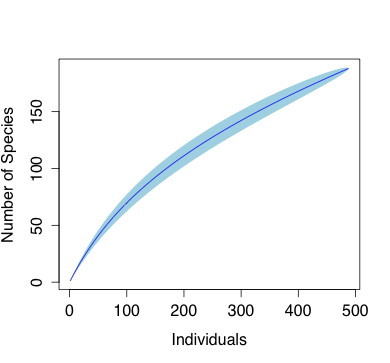
Fig.
Generalists vs. Specialists
We classified the forest understory into 6 types of microhabitats: saxicolous, epiphyllous, corticolous, terricolous, aquatic rocks (submerged occasionally during heavy rain), and lignicolous. The first five categories were almost exclusively found in the forest understory. The last category, lignicolous, consisted mainly of exposed dry fence posts on the northern boundary adjacent to a treeless deforested ranch. It is the only habitat that receives full sun (Table
|
No. Substrate |
Moss |
No. Specimens |
Liverwort |
No. Specimens |
Hornwort |
No. Specimens |
|
Types* |
Genera |
Collected |
Genera |
Collected |
Genera |
Collected |
|
6 |
0 |
0 |
1 |
97 |
0 |
0 |
|
5 |
0 |
0 |
1 |
39 |
0 |
0 |
|
4 |
8 |
30 |
5 |
232 |
0 |
0 |
|
3 |
9 |
73 |
14 |
140 |
1 |
14 |
|
2 |
17 |
66 |
14 |
74 |
2 |
4 |
|
1 |
40 |
225 |
40 |
89 |
0 |
0 |
|
Total |
74 |
394 |
75 |
671 |
3 |
18 |
*Substrate Types: Corticolous, epiphyllous, lignicolous, saxicolous, terricolous, and aquatic rocks,
Table
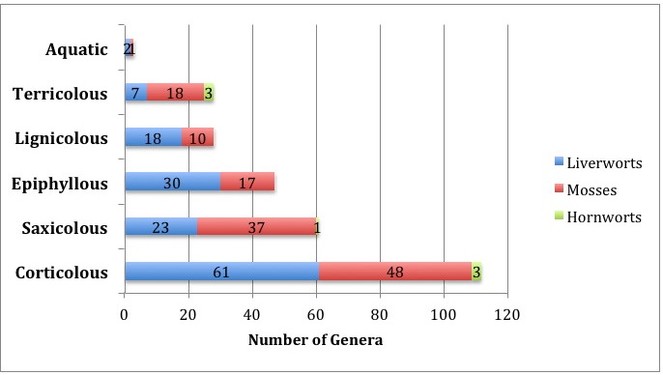
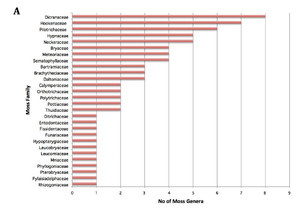
The corticolous bryophytes dominated with 112 genera collected off trunk, branches and twigs (Fig.
| Nectandra | Monteverde | Los Robles | Cocos Islands | Santa Elena Peninsula | |
| Citations |
|
|
|
|
|
| Exposure | Atlantic | Pacific | Pacific | Pacific | Pacific |
| Holdridge Life zones | premontane cloud forest | lower montane cloud forest | upper montane cloud forest | lowland and montane cloud forest | dry lowland & wet montane cloud forest |
| Period of Study | 2007-2010 | 1992-1994 | 1999-2000 | 1994 | 2003 |
| Location in Costa Rica |
|
|
|
|
|
| Elevation in m | 1100-1200 | 1550 | 2200-2500 | 0-600 | 0-700 |
| Survey Method | 4m x 10km trails | 4 x 1ha plots | 8m x 7km trails | 8 x 100m2 plots | Not given |
| Area Surveyed | 4 ha | 4 ha | 6 ha | 0.8 ha | Not given |
| Mean Annual Rainfall (m) | 3.5 | 2.5 | 2.8 | 6 | 1.5 |
| Mean Annual Temp °C | 20 | 18.8 | 11 | 25.5 | 28 |
| Collection height | ≤2m | lower canopy | Understorey and | Not Given | Not given |
| crown of 7 fallen trees | |||||
| Sampling | Haphazard | Random | Random | Quandrants 0.3m x 0.3m | Random |
| Forest type | Mature | Primary & secondary | Mature Oak | varied | varied |
| Total Moss Genera | 74 | 47 | 72 | 32 | 20 |
| Shared genera | 43 (91%) | 60 (83%) | 32 (100%) | 19 (95%) | |
| Different genera | 4 (9 %) | 12 (17%) | 0 (0%) | 1 (5%) | |
| Total Liverwort genera | 75 | 50 | 50 | 44 | 16 |
| Shared genera | 46 (92%) | 41 (83%) | 44 (100%) | 13 (81%) | |
| Different genera | 4 (8%) | 9 (17%) | 0 (0%) | 3 (19%) | |
| Total hornwort genera | 3 | 1 | 1 | 1 | 0 |
| Shared genera | 1 | 0 | 1 | ||
| Different genera | 0 | 1 | 0 |
Comparison of Five Locations from Different Life Zones
We made detailed comparisons of the bryological diversity, at the level of genera at Nectandra with 4 other locations previously mentioned in the Introduction. Table
Number of moss genera by family at the five sites. Each dot (•) = one shared genus; Each (o) = genus not at Nectandra.
| Mosses | Nectandra | Monteverde | Los Robles | Cocos | Santa Elena |
| Bartramiaceae | ••• | • | ••• | • | • |
| Brachytheciaceae | ••• | •• | •• | ||
| Bryaceae | •••• | ••• | •••• | • | •• |
| Calymperaceae | •• | • | • | •• | •• |
| Daltoniaceae | ••• | •• | ••• | ||
| Dicranaceae | •••••••• | ••••• | •••••• | •• | • |
| Ditrichaceae | • | ||||
| Entodontaceae | • | • | |||
| Fissidentaceae | • | • | • | • | • |
| Funariaceae | • | ||||
| Hookeriaceae | ••••••• | • | • | •• | |
| Hypnaceae | ••••• | •••• | •••• | ••• | • |
| Hypopterygiaceae | • | • | |||
| Leucobryaceae | • | • | •• | • | |
| Leucomiaceae | • | • | • | ||
| Meteoriaceae | •••• | •••••• | •••••• | •• | • |
| Mniaceae | • | • | |||
| Neckeraceae | ••••• | ••• | •• | • | |
| Orthotrichaceae | •• | ••••• | ••• | • | • |
| Phyllogoniaceae | • | • | • | ||
| Pilotrichaceae | •••••• | •••• | ••••• | ||
| Polytrichaceae | •• | •••• | |||
| Pottiaceae | •• | ••• | • | ••••• | |
| Pterobryaceae | • | • | • | • | |
| Pylaisiadelphaceae | • | ||||
| Rhizogoniaceae | • | ••• | •• | • | |
| Sematophyllaceae | •••• | ••• | ••• | ••••• | • |
| Thuidiaceae | •• | • | •• | • | |
| Adelotheciaceae | o | ||||
| Amblystegiaceae | o | o | |||
| Cryphaeaceae | o | ||||
| Hedwigiaceae | o | ||||
| Lembophyllaceae | o | o | |||
| Lepyrodontaceae | o | ||||
| Leucodontaceae | o | ||||
| Plagiotheciaceae | o | ||||
| Prionodontaceae | o | o | |||
| Racopilaceae | o | o | |||
| Regmatodontaceae | o | ||||
| Sphagnaceae | o | ||||
| Splachnaceae | o |
Number of liverwort genera by family of the five sites. Each dot (•) = one shared genus; Each (o) = genus not at Nectandra.
| Liverworts | Nectandra | Monteverde | Los Robles | Cocos | Santa Elena |
| Acrobolbaceae | • | • | • | ||
| Aneuraceae | •• | • | • | •• | |
| Aytoniaceae | •• | ||||
| Calypogeiaceae | • | • | • | • | |
| Cephaloziaceae | ••• | •• | •• | • | |
| Cephaloziellaceae | •• | • | • | ||
| Frullaniaceae | • | • | • | • | |
| Geocalycaceae | •• | •••• | •• | ||
| Herbertaceae | • | • | • | • | |
| Jamesoniellaceae | • | • | |||
| Lejeuneaceae | 40 | 26 | 23 | 28 | 11 |
| Lepicoleaceae | • | • | |||
| Lepidoziaceae | ••••• | ••••• | •••• | ••••• | |
| Lophocoleaceae | • | • | |||
| Marchantiaceae | • | • | |||
| Metzgeriaceae | • | • | • | ||
| Monocleaceae | • | • | |||
| Pallaviciniaceae | •• | •• | • | ||
| Pelliaceae | • | ||||
| Phycolepidoziaceae | • | ||||
| Plagiochilaceae | • | • | • | • | • |
| Pseudolepicoleaceae | •• | ||||
| Radulaceae | • | • | • | • | |
| Trichocoleaceae | • | • | • | ||
| Adelanthaceae | o | o | |||
| Balanthiopsaceae | o | ||||
| Fossombroniaceae | o | ||||
| Jubulaceae | o | o | |||
| Jungermanniaceae | o | ooo | |||
| Lepicoleaceae | o | ||||
| Porellaceae | o | ||||
| Ricciaceae | o | ||||
| Scapaniaceae | o | oo |
Discussion
Our inventory represents the first bryophyte collection at Nectandra and the first general bryophyte survey in a premontane cloud forest on the Atlantic slope of Costa Rica. Geologically, Nectandra and Monteverde are both located on the same volcanic range, the Cordillera Tilarán, within 50km of each other. Floristically, the two reserves share the same principal tree families, Lauraceae and Rubiaceae, as well as a whole host of terrestrial herbs and epiphytes (
Species diversity
From the accessions that we can determine with confidence, a total of 189 species were identified. This current number of species is far from complete, as suggested by increasing slope of the species accumulation curve and the discrepancy from the projected 213-412 species in Fig. 2. The remaining species are possibly among the undetermined specimens, or unexplored habitats, such as canopy or fully submerged habitat.
Generic density
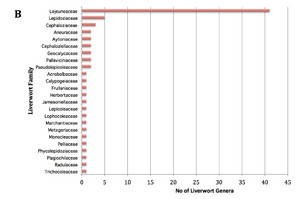
For practical reasons, we concentrated on the bryophytes accessible without climbing and included living as well as fallen trees. We did not estimate the proportion of specimens associated with freshly fallen trees. Our survey yielded a total of 152 genera of mosses, liverworts and hornworts at Nectandra, with a comparable number of hepatic and moss genera, although the number of genera per family differed markedly for the two groups (Fig.
Mosses compared to hepatics
While there was an almost equal number of genera collected for mosses and liverworts, the ratios of specimens per genus was not — 394/74 for mosses vs. 671/75 for liverworts. This difference can be accounted quantitatively and qualitatively by Lejeuneaceae. Not only were there 40 genera in that single family, 68 % of the genera were generalists, growing on 2 of the 6 substrates tested, hence the large number of total liverwort specimens encountered. At the same time that there was a higher generic diversity in Lejeuneaceae, the number of individuals per genus in this family is lower — 7.5 compared to the average 10.9 for liverworts in the remaining genera and 10.7 for mosses in general. Given that we employed haphazard floristic sampling, the number of specimens should reflect more or less the abundance of bryophytes in the garden. Such a difference in the specimen numbers of mosses and liverworts highlights the tropical nature of bryophyte distribution, in which liverworts tend to be more common than they do in the temperate zone (
Multi-site comparison
Our survey at Nectandra yielded 152 genera compared to 123 for Los Robles, 98 reported for Monteverde, 77 for Cocos and 36 for Santa Elena Peninsula. Given that the methodology and the surveyors differed at each site, it is not possible to make statistically meaningful comparisons. Nevertheless, it is useful to know the outcome of a coarse comparison among the five sites (Tables
Conclusions
Despite its partial account, our inventory of bryophytes at Nectandra yield the highest number of genera in the area-based studies in Costa Rica to date. The data here will hopefully contribute to a growing database and stimulate further floristic and ecological studies.
Acknowledgements
This work was co-funded by Dr. Daniel H. Norris and the Nectandra Institute. Professor S. Robbert Gradstein provided invaluable help in the identification of some of our samples. We thank Freddy Castillo for his specimen contribution and valuable on-site logistic assistance for the fieldwork at the Nectandra Cloud Forest Reserve.
Hosting institution
Dr. Daniel H. Norris' three bryophyte surveys and the 2010 Jepson Herbarium Tropical Bryophyte Workshop were hosted by Nectandra Cloud Forest Reserve, Costa Rica. This work was done under SINAC collecting permit 034-2010-SINAC.
Author contributions
DHN was the principal collector. EK and ETL collated the information on the specimens and prepared the manuscript. AR, DL, DA, FR, TAC, NF, MT, NP were participants of the 2010 Jepson Tropical Bryophyte Workshop who contributed specimens and discussion for this report.
References
- Hydrometeorology of tropical montane cloud forest: Emerging patterns.Hydrol. Process25:465‑498. https://doi.org/10.1002/hyp.7974
- Clark KL, Lawton RO, Butler PR (2000) The physical environment. In: Nadkarni NM, Wheelwright NT (Eds) Monteverde: Ecology and Conservation of a tropical Cloud Forest.Oxford University Press,New York, New York,15-19pp.
- Physical environment, in Monteverde: Ecology and Conservation of a Tropical Cloud Forest - 2014 Updated Chapters. http://digitalcommons.bowdoin.edu/scholars-bookshelf/5
- Retention of inorganic nitrogen by epiphytic bryophytes in a tropical montane forest.C. Biotropica37(3):328‑336. https://doi.org/10.1111/j.1744-7429.2005.00043.x
- Atmospheric deposition and net retention of ions by the canopy of a tropical montane forest, Monteverde, Costa Rica.J trop ecol14:27‑45. https://doi.org/10.1017/S0266467498000030
- Nutrient release from epiphytic bryophytes in tropical montane rain forest (Guadeloupe).J Bot69(10):2122‑2129.
- Bryophytes of Cocos Island, Costa Rica: diversity, biogeography and ecology.Rev biol trop47:1‑17.
- Bryophytes of the Santa Elena Peninsula and Islas Mucielago, Guanacaste, Costa Rica, with special attention to neotropical dry forest habitats.Lankesteriana5(1):53‑6.
- Bryophyte Biology.Cambridge University Press,Cambridge,565pp.
- The incidence and implications of clouds for cloud forest water relations.Ecology Letters16:307‑314. https://doi.org/10.1111/ele.12039
- A classification of Lejeuneaceae (Marchantiophyta) based on molecular and morphological evidence.Phytotaxa100:6‑20. https://doi.org/10.11646/phytotaxa.100.1.2
- Guide to the Bryophytes of Tropical America.New York Botanical Garden Press,Bronx, NY,573pp.
- Diversity and habitat differentiation of mosses and liverworts in the cloud forest of Monteverde, Costa Rica.Caldasia23(1):203‑212.
- Morphological and anatomical features associated with epiphytism among the pleurocarpous mosses — one basis for further research on adaptations and their evolution.Journal of Bryology34:79‑100. https://doi.org/10.1179/1743282011Y.0000000049
- Life zone ecology.Tropical Science Center,San Jose, Costa Rica,149pp.
- Bryophyte diversity, microhabitat differentiation and distribution of life forms in Costa Rican upper montane Quercus forest.The Bryologist105(3):334‑348. https://doi.org/10.1639/0007-2745(2002)105[0334:BDMDAD]2.0.CO;2
- Lawton RO, Nair US, Ray D, Pounds JA, Welch RM (2010) Quantitative measures of immersion in cloud and the biogeography of cloud forest. In: Bruijnzeel LA., Scatena FN, Hamilton LS (Eds) Tropical Montane Cloud Forests – Science for Conservation and Management.
- Nitrogen fixation in bryophytes, lichens and decaying wood along a soil-age gradient in Hawaiian montane rain forest.Biotropica35(1):12‑19. https://doi.org/10.1646/0006-3606(2003)035[0012:NFIBLA]2.0.CO;2
- Epiphytic bryophytes of Monteverde, Costa Rica.Trop Bryo20:63‑70.
- Ecology and Conservation of a tropical Cloud Forest, Monteverde, Costa Rica.Oxford University Press,New York, New York,481pp.
- Biogeography of tropical montane cloud forests. Part II: Mapping of orographic cloud immersion.J App Met Clim47:2183‑2197. https://doi.org/10.1175/2007JAMC1819.1
- vegan: Community Ecology Package. R package.version 2.4-0. Release date:2016-9-07. URL: http://CRAN.R-project.org/package=vegan
- The hydroclimatic and ecophysiological basis of cloud forest distributions under current and projected climates.Ann Bot113(6):909‑920. https://doi.org/10.1093/aob/mcu060
- Pócs T (1982) Tropical Forest Bryophytes. In: Smith AJE (Ed.) Bryophyte ecology.Springer NetherlandsURL: : http://www.springerlink.com/index/10.1007/978-94-009-5891-3_3.
- Pollawatn R (2008) Systematic treatment of Sematophyllaceae (Musci) in Thailand.Rheinischen-Friedrich-Wilhelms-Universität Bonn,14 MBpp. URL: http://hss.ulb.uni-bonn.de/2008/1449/1449.htm
- Synbiotic fungal associations in “lower” land plants.Philosophical Transactions of the Royal Society of London, Series B-Biological Sciences355:815‑830. https://doi.org/10.1098/rstb.2000.0617
- The role of bryophytes in carbon and nitrogen cycling.The Bryologist10(63):395‑409.
Supplementary materials
List of 1083 bryophyte specimens collected at Nectandra Cloud Forest Reserve (2007-2010) with accompanying habitat and collection information. These specimens were identified to genus level
List of 488 of bryophyte specimens collected at Nectandra (2007-2010) with full identification