|
Biodiversity Data Journal :
Single Taxon Treatment
|
|
Corresponding author: Carolyn Trietsch (cut162@psu.edu)
Academic editor: Jose Fernandez-Triana
Received: 30 Nov 2017 | Accepted: 22 Jan 2018 | Published: 30 Jan 2018
© 2018 Carolyn Trietsch, István Mikó, David Notton, Andrew Deans
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Trietsch C, Mikó I, Notton D, Deans A (2018) Unique extrication structure in a new megaspilid, Dendrocerus scutellaris Trietsch & Mikó (Hymenoptera: Megaspilidae). Biodiversity Data Journal 6: e22676. https://doi.org/10.3897/BDJ.6.e22676
|
|
Abstract
Background
A new species, Dendrocerus scutellaris Trietsch & Mikó (Hymenoptera: Megaspilidae), is described here from male and female specimens captured in Costa Rica. This species is the only known ceraphronoid wasp with a straight mandibular surface and raised dorsal projections on the scutellum, called the mesoscutellar comb. It is hypothesised that the function of the mesoscutellar comb is to aid the emergence of the adult from the host, especially since the mandibles lack a pointed surface to tear open the pupal case. The authors also provide phenotypic data in a semantic form to facilitate data integration and accessibility across taxa and provide an updated phenotype bank of morphological characters for megaspilid taxonomic treatments. In updating this phenotype bank, the authors continue to make taxonomic data accessible to future systematic efforts focusing on Ceraphronoidea.
New information
A new species, Dendrocerus scutellaris (Hymenoptera: Megaspilidae) Trietsch & Mikó, is described from both male and female specimens captured in Costa Rica.
Keywords
Ceraphronoidea, morphology, systematics, taxonomy, eclosion
Introduction
Ceraphronoidea is a relatively small superfamily of parasitoid wasps with a worldwide distribution (
Eclosion is the adult emergence from the pupal case in holometabolous insects. In most holometabolous insects, the tearing of the pupal case is achieved by the movement of the insect and the increased hemolymph pressure caused by muscle contractions (
In Hymenoptera, wood-boring families have been observed to have specialised structures for extricating themselves from the pupal chambers inside wood where they develop (
Here, a new species is described of Dendrocerus captured in Costa Rica characterised by the presence of a straight mandibular edge and the mesoscutellar comb, which could aid in emergence. These two structures have never before been recorded in Ceraphronoidea and are discussed here for the first time.
Materials and methods
Point-mounted specimens were borrowed from the Natural History Museum (NHMUK) in London, United Kingdom. Specimen data is provided in Suppl. material
Point-mounted and glycerine-dissected specimens were examined using an Olympus SZX16 stereomicroscope with an Olympus SDF PLAPO 1XF objective (115×) and an Olympus SDF PLAPO 2XPFC objective (230× magnification). Blue-Tac (Bostik, Inc., Wauwatosa, Wisconsin, USA) and molding clay (Sculpey, Polyform Products Company, Elk Grove Village, Illinois, USA) was used to stabilise specimens during imaging and observation. Stacks of bright field images were taken manually on an Olympus CX41 microscope with a Canon EOS 70D camera attached. Images were subsequently aligned and stacked using Zerene Stacker Version 1.04 Build T201706041920. Figures were created in Adobe Photoshop elements Version 3.1.
To prepare specimens for male genitalia dissection, metasomata were removed from point-mounted specimens and cleared with 35% H2O2 (Alfa Aesar) for 24 hours, then moved to 5% acetic acid (Distilled White Vinegar, Great Value) for 24 hours and subsequently moved to glycerol for dissection and short-term storage. Dissections were performed with #5 forceps (Rubis 5A-SA, Bioquip) and #2 insect pins (BioQuip). Male genitalia were then mounted between 1.5 mm thick, 24×50 mm cover glasses and imaged using an Olympus FV10i confocal laser scanning microscope. Following the methods of
For the descriptions of male and female specimens, morphological characters (following
Semantic statements were generated in Protégé Version 5.0 beta-15 following the methods of
All figures, media files, protocols, semantic statements and supplementary files are available on figshare at https://figshare.com/projects/Unique_extrication_structure_in_a_new_megaspilid_Dendrocerus_scutellaris_Hymenoptera_Megaspilidae_Trietsch_and_Mik_/27007. The taxonomic treatment file generated from MX (Suppl. material
Taxon treatment
Dendrocerus scutellaris , sp. n.
-
scientificName: Dendrocerus scutellaris; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Megaspilidae; taxonRank:species; genus:Dendrocerus; specificEpithet:scutellaris; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; verbatimLocality:COSTA RICA: Guanacaste Pr.: Santa Rosa N. P.: 300m Bosque San Emilio; verbatimElevation:300m; eventDate:1985-04-27/5-11; verbatimEventDate:27.iv-11.v.1985; eventRemarks:sample SE.6.C. BMNH(E) 2008-87; individualCount:1; sex:male; lifeStage:adult; catalogNumber:NHMUK010812028; recordedBy:D Janzen & I. Gauld; identifiedBy:Carolyn Trietsch; dateIdentified:2017; language:en; institutionID: ; institutionCode:NHMUK; collectionCode:Insects; basisOfRecord:PreservedSpecimen; occurrenceID:
-
scientificName: Dendrocerus scutellaris; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Megaspilidae; taxonRank:species; genus:Dendrocerus; specificEpithet:scutellaris; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; verbatimLocality:COSTA RICA: Guanacaste Pr.: Santa Rosa N. P.: 300m Bosque San Emilio; verbatimElevation:300m; eventDate:1985-10-5/26; verbatimEventDate:5-26.x.1985; eventRemarks:sample SE.6.C. BMNH(E) 2008-87; individualCount:1; sex:female; lifeStage:adult; catalogNumber:NHMUK010812044; recordedBy:D Janzen & I. Gauld; identifiedBy:Carolyn Trietsch; dateIdentified:2017; language:en; institutionID: ; institutionCode:NHMUK; collectionCode:Insects; basisOfRecord:PreservedSpecimen; occurrenceID:
-
scientificName: Dendrocerus scutellaris; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Megaspilidae; taxonRank:species; genus:Dendrocerus; specificEpithet:scutellaris; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; verbatimLocality:COSTA RICA: Guanacaste Pr.: Santa Rosa N. P.: 300m Bosque San Emilio; verbatimElevation:300m; eventDate:1985-07-5/8-3; verbatimEventDate:13.vii-3.viii.1985; eventRemarks:sample SE.6.C. BMNH(E) 2008-87; individualCount:1; sex:female; lifeStage:adult; catalogNumber:NHMUK010812045; recordedBy:D Janzen & I. Gauld; identifiedBy:Carolyn Trietsch; dateIdentified:2017; language:en; institutionID: ; institutionCode:PSUC; collectionCode:Insects; basisOfRecord:PreservedSpecimen; occurrenceID:
-
scientificName: Dendrocerus scutellaris; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Megaspilidae; taxonRank:species; genus:Dendrocerus; specificEpithet:scutellaris; country:Costa Rica; countryCode:CR; stateProvince:Guanacaste; verbatimLocality:COSTA RICA: Guanacaste Pr.: Santa Rosa N. P.: 300m Bosque San Emilio; verbatimElevation:300m; eventDate:1985-07-5/8-3; verbatimEventDate:13.vii-3.viii.1985; eventRemarks:sample SE.6.C. BMNH(E) 2008-87; individualCount:1; sex:male; lifeStage:adult; catalogNumber:NHMUK010812030; recordedBy:D Janzen & I. Gauld; identifiedBy:Carolyn Trietsch; dateIdentified:2017; language:en; institutionID: ; institutionCode:NHMUK; collectionCode:Insects; basisOfRecord:PreservedSpecimen; occurrenceID:
Description
Body length universal: 2.6-2.7 mm.
Colouration: Colour hue pattern: head and mesosoma black; metasoma, mouthparts, legs and scape except for the basal part dark brown; base of scape light brown. Colour intensity pattern: proximal part of scape lighter than the rest of the scape.
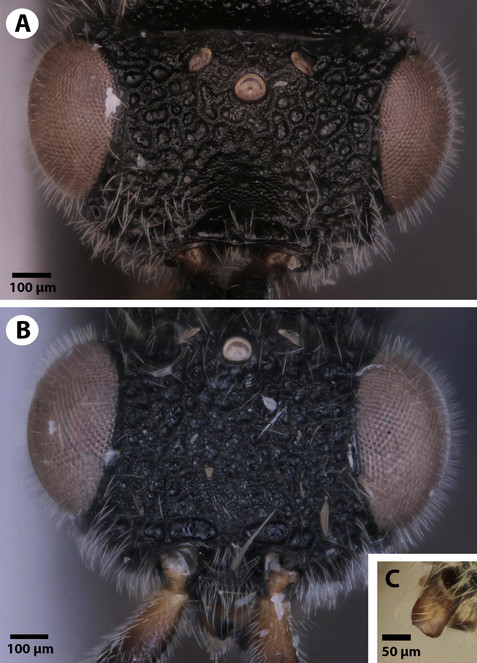
Head: Cephalic size (csb): mean: 750-1100 μm. Head height (lateral view) vs. eye height (anterior view): HH:EHf=1.25-1.75. Head height vs. head length: HH:HL=1.2-1.5. Head width vs. interorbital space: HW:IOS=1.6-1.9. Head width vs. head height: HW:HH=1.5-2.0. Male ocular ocellar line vs. lateral ocellar line: OOL:LOL=2.1-2.6. Male ocular ocellar line vs. posterior ocellar line: OOL:POL=0.95-1.0. Female ocular ocellar line vs. lateral ocellar line: OOL 1.6–2.5 x as long as LOL. Anterior ocellar fovea shape: fovea not extended ventrally into facial sulcus. Occipital carina sculpture: crenulate. Median flange of occipital carina count: absent. Preoccipital carina count: present. Preoccipital lunula count: present. Preoccipital furrow count: present. Preoccipital furrow anterior end: preoccipital furrow ends inside ocellar triangle. Dorsal margin of occipital carina vs. dorsal margin of lateral ocellus in lateral view: occipital carina is ventral to lateral ocellus in lateral view. Transverse scutes on upper face count: absent. Rugose region on upper face count: present. Rugose sculpturing on head and mesosoma count: present. Facial pit count: facial pit present. Intertorular carina count: present. Ventral margin of antennal rim vs. dorsal margin of clypeus: not adjacent. Median region of intertorular area shape: concave. Subtorular carina count: present. Torulo-clypeal carina count: present. Supraclypeal depression count: present. Supraclypeal depression structure: absent medially, represented by two grooves laterally of facial pit. Antennal scrobe count: absent. Mandibular tooth count: 1. Mandibular lancea count: absent. Distal edge of mandible: flat.
Antennae: Male flagellomeres shape: branched. Male scape length vs combined length of F1+F2: longer or equal. 6th male flagellomere length vs. width, “sensillar” view: elongate, more than 2x as long as wide. Male flagellomere branches count: 7 branches ; 8 branches . Branch of male flagellomere 5 length compared to flagellomere 6: longer than length of flagellomere 6. Branch of male F5 length vs. length of male F5: longer than length of flagellomere 5. Male F6 length vs. combined length of F7+F8: shorter than length of flagellomere 7+8. Sensillar patch of the male flagellomere pattern: F7-F9. Basal resilin-rich area of male antennal branches count: absent. Female F1 length vs. pedicel length: 1.0-1.2. Female ninth flagellomere length: F9 less than F7+F8.
Mesosoma and Metasoma: Ventrolateral invagination of the pronotum count: present. Notaulus posterior end location: adjacent to transscutal articulation. Speculum ventral limit: not extending ventrally of pleural pit line. Mesoscutellar comb count: present. Mesoscutal length vs. anterior mesoscutal width: MscL/AscW=1.2–2.0. Anterior mesoscutal width vs. posterior mesoscutal width: AscW/PscW=0.7-0.9. Median mesoscutal sulcus posterior end: adjacent to transscutal articulation. Axillular carina count: absent. Scutoscutellar sulcus vs. transscutal articulation: adjacent. Mesometapleural sulcus count: present. Metapleural carina count: present. Anteromedian projection of the metanoto-propodeo-metapecto-mesopectal complex count: present. Anteromedian projection of the metanoto-propodeo-metapecto-mesopectal complex shape: Bifurcated.
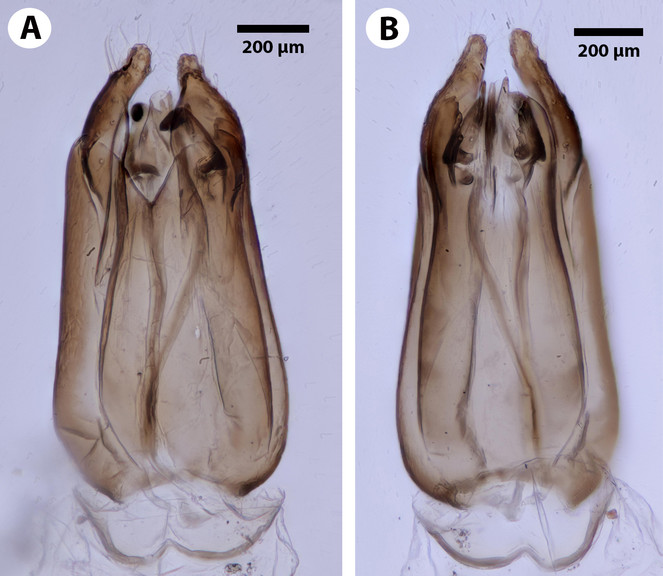
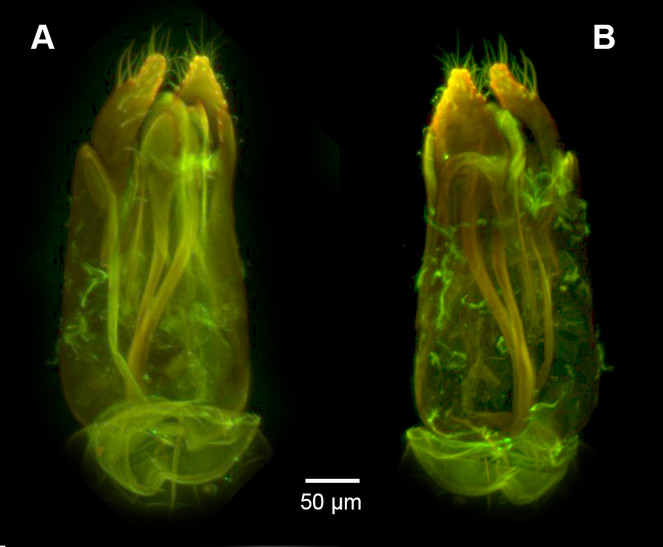
Male Genitalia: Distal margin of male S9 shape: convex. Proximolateral corner of male S9 shape: blunt. Proximodorsal notch of cupula count: absent. Gonostyle/volsella complex proximodorsal margin shape: with deep concavity medially. Submedian conjunctiva on distoventral margin of gonostyle/volsella complex: length (range of fusion of parossiculus/parossiculus complex from gonostipes): more than 4/5. Apical parossiculal seta number: one. Dorsal apodeme of penisvalva count: absent. Distal projection of the penisvalva count: absent. Sensillar plate of the aedeagus shape: distinctly less than half as wide as the male genitalia. Distal projection of the parossiculus count: present. Dorsomedian conjunctiva of the gonostyle-volsella complex count: absent. Cupula length vs. gonostyle-volsella complex length: cupula less than 1/2 the length of gonostyle-volsella complex in lateral view. Parossiculus count (parossiculus and gonostipes fusion): present (not fused with the gonostipes). Distoventral submedian corner of the cupula count: absent. Harpe length: harpe shorter than gonostipes in lateral view.
Diagnosis
Dendrocerus scutellaris (Figs



Etymology
This species is named for the presence of the mesoscutellar comb, which is unique to this species and is not found in any other known ceraphronoid species.
Distribution
This species is only known from Costa Rica.
Discussion
Dendrocerus scutellaris belongs to the halidayi species-group, which is characterised by the presences of flabellate antennae in males (
D. scutellaris is unique amongst members of the halidayi species-group in that, while other species have up to six fully formed branches on the flagellomeres (
Dendrocerus scutellaris is distinguished from all other ceraphronoid species by the presence of a straight mandibular surface (Fig.
Acknowledgements
The authors thank Missy Hazen for her expertise and assistance with CLSM at the Penn State Microscopy and Cytometry Facility (University Park, PA). The authors would also like to thank the reviewers, Simon van Noort and Aniruddha Mitra, for their insightful comments and help in improving the manuscript. This material is based upon work supported by the U. S. National Science Foundation, under Grant Numbers DBI-1356381 and DEB-1353252. Any opinions, findings and conclusions or recommendations expressed in this material are those of the authors and do not necessarily reflect the views of the National Science Foundation.
References
- New species of Platygastridae (Hymenoptera) from India which parasitise pests of mango, particularly Procontarinia spp. (Diptera: Cecidomyiidae).Bulletin of Entomological Research74:549‑557. https://doi.org/10.1017/S0007485300013924
- Presence-absence reasoning for evolutionary phenotypes.arXiv preprint arXiv:1410.3862.
- A semantic model for species description applied to the Ensign wasps (Hymenoptera: Evaniidae) of New Caledonia.Systematic Biology62(5):639‑659. https://doi.org/10.1093/sysbio/syt028
- Dendrocerus mexicali (Hymenoptera, Ceraphronoidea, Megaspilidae): Novel antennal morphology, first description of female, and expansion of known range into the U.S.ZooKeys569:53‑69. https://doi.org/10.3897/zookeys.569.6629
- Host handling and specificity of the hyperparasitoid wasp, Dendrocerus carpenteri (Curtis) (Hym. Megaspilidae): importance of host age and species.Journal of Applied Entomology123(2):83‑91. https://doi.org/10.1046/j.1439-0418.1999.00322.x
- A review of the West Indian moths of the family Psychidae with descriptions of new taxa and immature stages.Smithsonian Contributions to Zoology188:1‑66. https://doi.org/10.5479/si.00810282.188
- Time to change how we describe biodiversity.Trends in Ecology & Evolution27(2):78‑84. https://doi.org/10.1016/j.tree.2011.11.007
- A propos du genre Dendrocerus Ratzeburg, 1852. Les espèces du group "penmaricus" (Hymenoptera Ceraphronoidea Megaspilidae).Bulletin et Annales de la Societe Royale Belge d'Entomologie (Belgium)131:349‑382.
- Révision des Dendrocerus du groupe «halidayi» (Hym. Ceraphronoidea Megaspilidae).Belgian Journal of Entomology1:169‑275.
- The Insects: An Outline of Entomology.4th.Wiley-Blackwell
- Revision of world species of the genus Heptascelio Kieffer (Hymenoptera: Platygastroidea, Platygastridae).Zootaxa1776:1‑51.
- Catalog of the systematic literature of the superfamily Ceraphronoidea (Hymenoptera).Contributions of the American Entomological Institute33:1-149.‑1-149..
- Facultative gregarious development in a solitary parasitoid wasp, Dendrocerus carpenteri: larvae may share nutritional resources.Entomologia Experimentalis et Applicata157(2):170‑180. https://doi.org/10.1111/eea.12352
- Resource rivalry between brood mates of a facultative gregarious parasitoid Dendrocerus carpenteri: Resource rivalry in parasitoid larvae.Physiological Entomology42(1):65‑72. https://doi.org/10.1111/phen.12167
- What is fluorescing?Hamuli4:19‑22. URL: http://www.hymenopterists.org/newsletters/hamuli/HamuliVol4Issue2.pdf
- Folding wings like a cockroach: A review of transverse wing folding ensign wasps (Hymenoptera: Evaniidae: Afrevania and Trissevania).PLoS ONE9(5):e94056. https://doi.org/10.1371/journal.pone.0094056
- Generating semantic phenotypes.figsharehttps://doi.org/10.6084/m9.figshare.1314904.v1
- Host discrimination and ovicide by aphid hyperparasitoids Asaphes suspensus (Hymenoptera: Pteromalidae) and Dendrocerus carpenteri (Hymenoptera: Megaspilidae).Applied Entomology and Zoology51(4):609‑614. https://doi.org/10.1007/s13355-016-0437-0
- The mode of extrication of silkworm moths from their cocoons.The American Naturalist12(6):379‑383. https://doi.org/10.1086/272118
- A new species of Dendrocerus (Hymenoptera, Megaspilidae) from southern Brazil.Zookeys425:51‑57. https://doi.org/10.3897/zookeys.425.7454
- A review of the Afrotropical Rhyssinae (Hymenoptera: Ichneumonidae) with the descriptions of five new species.European Journal of Taxonomy91[InEnglish]. https://doi.org/10.5852/ejt.2014.91
- Fiji: an open-source platform for biological-image analysis.Nature Methods9(7):676‑682. https://doi.org/10.1038/nmeth.2019
- The ImageJ ecosystem: An open platform for biomedical image analysis.Molecular Reproduction and Development82(7-8):518‑529. https://doi.org/10.1002/mrd.22489
- Foraging for mates in the hyperparasitic wasp, Dendrocerus carpenteri: impact of unfavourable weather conditions and parasitoid age.Oecologia119(1):73‑80. https://doi.org/10.1007/s004420050762
- Redescription of Conostigmus albovarius Dodd, 1915 (Hymenoptera, Megaspilidae), a metallic ceraphronoid, with the first description of males.Journal of Hymenoptera Research46:137‑150. https://doi.org/10.3897/JHR.46.5534
- Into the wood and back: morphological adaptations to the wood-boring parasitoid lifestyle in adult aulacid wasps (Hymenoptera: Aulacidae).Journal of Hymenoptera Research19(2):244‑258.
- Revision of the Afrotropical Mayrellinae (Cynipoidea: Liopteridae), with the first record of Paramblynotus from Madagascar.Journal of Hymenoptera Research31:1‑64. [InEnglish]. https://doi.org/10.3897/jhr.31.4072
- Per arborem ad astra: Morphological adaptations to exploiting the woody habitat in the early evolution of Hymenoptera.Arthropod Structure & Development40(1):2‑20. https://doi.org/10.1016/j.asd.2010.10.001
- Ontobee: A linked data server and browser for ontology terms.Proceedings of the 2nd International Conference on Biomedical Ontologies (ICBO), July 28-30, 2011, Buffalo, NY, USA.279-281.‑279-281..
- A gross anatomy ontology for Hymenoptera.PloS ONE5(12):e15991. https://doi.org/10.1371/journal.pone.0015991
- Eclosion behavior in tsetse (Diptera: Glossinidae): extrication from the puparium and expansion of the adult.Journal of Insect Behavior5(5):657‑668. https://doi.org/10.1007/BF01048011
Supplementary materials
A table listing all of the specimens used in this study and their associated locality and repository information.
The taxonomic treatment file generated from MX used to write semantic statements.
The file containing all of the semantic statement phenotype annotations.