|
Biodiversity Data Journal :
Taxonomic Paper
|
|
Corresponding author: Hannes Baur (hannes.baur@nmbe.ch)
Academic editor: Matthew Yoder
Received: 24 Jun 2018 | Accepted: 31 Oct 2018 | Published: 14 Nov 2018
© 2018 Fabian Klimmek, Hannes Baur
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Klimmek F, Baur H (2018) An interactive key to Central European species of the Pteromalus albipennis species group and other species of the genus (Hymenoptera: Chalcidoidea: Pteromalidae), with the description of a new species. Biodiversity Data Journal 6: e27722. https://doi.org/10.3897/BDJ.6.e27722
|
|
Abstract
Background
Parasitoid wasps of the genus Pteromalus play an important role in biological pest control, however, the genus includes a large number of cryptic species, which makes reliable identification difficult. The latest identification key dates back to Graham (1969) and since then many new species have been described and nomenclatural changes proposed.
New information
Here we present an interactive and fully illustrated identification key in Xper3 for 27 species of the Pteromalus albipennis species group as well as for 18 similar species. In addition to qualitative traits, a large set of body measurements is incorporated in the key. We also explored a new set of qualitative features on the propodeum and metasternum. During field work, a new species of the P. albipennis species group, P. capito Baur sp. n., could be reared from flower heads of Asteraceae, which is described here. It looks very similar to P. albipennis and P. cingulipes, however, several qualitative characters and body ratios distinguish it clearly from the most similar species.
Keywords
Diptera, microtomy, paraspiracular inclination, Pteromalinae, P. achillei, P. caudiger, P. cingulipes, P. eudecipiens, P. intermedius, P. patro, P. puparum, P. temporalis, shape principal component analysis, taxonomy, Tephritidae
Introduction
Recently developed software for the creation of biological identification keys like Xper3 (
Here we created an interactive identification key in Xper3 for European Pteromalus with special focus on the P. albipennis species group. The latest key for the identification of Pteromalus is a monograph on the Pteromalidae of North-Western Europe (
Parasitoid wasps of the genus Pteromalus belong to the family Pteromalidae (Insecta: Hymenoptera: Chalcidoidea). The genus contains 371 known species in Europe (
Materials and methods
Creation of the key
The key was created in Xper3 version 1.4.0. Pictures could be uploaded as JPEGs and are hosted on a free Dropbox account. The original TIF-files are deposited on zenodo.org. Descriptors were ranked with the built-in ranking system of Xper3 (rank 1 = little important, 5 = highly important) according to their distinct nature and relative simplicity to study. Measurements, which were taken for all species, were read in as quantitative data with fixed ranges. Completeness of the database was checked using the inbuilt tools for analysis of the data.
Use of Xper3
As the official user manual of Xper3 (
The user of Xper3 has several tabs to edit the descriptive data in the software. In the “Items” section all taxa are listed and can be provided with additional detailed information (e.g. distribution, first author, habitat, indicator values etc.) and with illustrating material like pictures, movies, sound files or any other file (e.g. maps, text files). In addition, new taxa can be added and edited in the same way as already existing taxa. The “Descriptive model” section lists all descriptors and can also be edited as described for the “Items” section. States for each descriptor can be edited or new ones can be added.
Xper3 has the possibility to weight the descriptors. This is a great way to simplify and speed up the identification process, allowing quick and reliable results. For instance, problematic (e.g. shape of propodeal foramen) or rather difficult (e.g. degree of paraspiracular inclination) characters can be weighted less, which reduces the chances for users to deal with them already at the beginning of the identification process. However, there is no guarantee that such characters will never appear in the top part of the list.
The section “Description” offers the possibility to assign each descriptor the respective states for each item.
Xper3 comes with some diagnostic tools to check the completeness of the database or to analyse the data. A view of the complete description matrix with the possibility to search for undescribed items and the revision of these is a great help to make sure that each taxon is completely described.
Furthermore, it is possible to select various items or assign them to groups for comparison. The output is a matrix, which also shows all descriptor states of every included item. In addition, the states for each descriptor, where the items differ or are congruent, are displayed separately.
The tool “Checkbase” is an automated search for various errors in the database, for instance items with identical descriptions, undescribed or inapplicable descriptors and descriptors described as unknown. A direct link to these errors provides a handy means for quick and easy revision. Finally, it is possible to generate different kinds of keys. Besides the interactive version, flat or tree-shaped dichotomous versions can be generated in various output formats (HTML, PDF, SDD, Zip archive etc.).
To work with the key, one selects any character which seems conspicuous or unique to the specimen and decides which state is appropriate. Of course, it is recommended to start with the top characters, as they have been weighted higher by the author for good reason. Once a selection is made, the software will exclude every taxon which does not fit this set of states and give a list of the remaining possible names. If more than one name remains, more characters have to be studied. If only one name remains, this should normally be the name one is looking for. This can be checked by comparing the specimen with all the pictures linked to this taxon and the complete diagnosis. The section “History” provides a helpful means to look up the identification process and correct potential mismatches without resetting the complete selection.
Validity of the key
The taxa to be included in the key were based on a list of valid European species for the genus Pteromalus from the Universal Chalcidoidea Database (
Of the remaining 73 names, we included 28 species belonging to the P. albipennis species group (sensu
Of the remaining 47 names, we included 9 species from 8 species groups (sensu
List of species of the Pteromalus albipennis species group and other species included in the key.
|
Group |
Species |
|
P. albipennis species group (29 species) |
aartseni (Gijswijt, 1972) |
|
achillei Janzon, 1984 |
|
|
albipennis Walker, 1835 |
|
|
ametrus Graham, 1981 |
|
|
berylli Walker, 1835 |
|
|
brachygaster (Graham, 1969) |
|
|
capito Baur sp.n. |
|
|
cardui (Erdös, 1953) |
|
|
caudiger (Graham, 1969) |
|
|
conformis (Graham, 1969) |
|
|
costulata Gijswijt, 1999 |
|
|
cingulipes Walker, 1835 |
|
|
elevatus (Walker, 1834) |
|
|
eudecipiens Özdikmen, 2011 [P. decipiens (Graham, 1969)] |
|
|
inclytus Förster, 1841 |
|
|
integer Walker, 1872 |
|
|
intermedius (Walker, 1834) |
|
|
lactucae (Szelényi & Erdös, 1953) |
|
|
musaeus Walker, 1844 |
|
|
myopitae (Graham, 1969) |
|
|
parietinae (Graham, 1969) |
|
|
patro Walker, 1848 |
|
|
pilosellae Janzon, 1984 |
|
|
scandiae (Graham, 1969) |
|
|
solidaginis Graham & Gijswijt, 1991 |
|
|
speculifer Graham, 1981 |
|
|
temporalis (Graham, 1969) |
|
|
tibiellus Zetterstedt, 1838 |
|
|
tripolii (Graham, 1969) |
|
|
Other Pteromalus (18 species) |
altus (Walker, 1834) |
|
bedeguaris (Thomson, 1878) |
|
|
bifoveolatus Förster, 1861 |
|
|
briani Baur, 2015 |
|
|
chlorospilus (Walker, 1834) |
|
|
chrysos Walker, 1836 |
|
|
crassinervis (Thomson, 1878) |
|
|
cyniphidis (Linnaeus, 1758) |
|
|
elatus Förster, 1841 |
|
|
hieracii (Thomson, 1878) |
|
|
janstai Baur, 2015 |
|
|
ochrocerus (Thomson, 1878) |
|
|
platyphilus Walker, 1874 |
|
|
puparum (Linnaeus, 1758) |
|
|
semotus (Walker, 1834) |
|
|
sequester Walker, 1835 |
|
|
vibulenus (Walker, 1839) |
|
|
vopiscus Walker, 1839 |
If the key leads to species names representing whole species groups (e.g. altus, sequester etc.), the resulting identification has to be treated with caution. In this case, we refer to (Graham 1969) for a sound identification.
Nomenclature and classification of Pteromalidae follow
Material examined
All specimens relevant to this work are listed in appendix A. Examined specimens are deposited in the following collections (abbreviations according to
Preparation
Most characters were studied on dried and card mounted specimens (
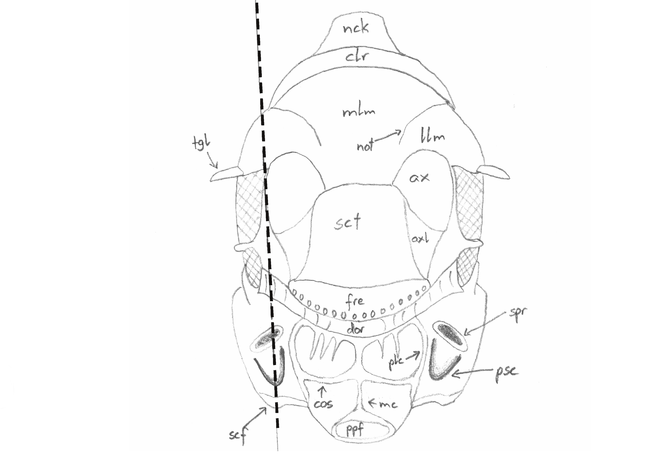



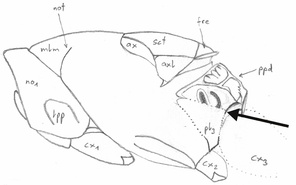
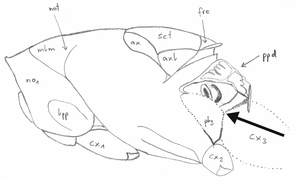



Measurements
Characters were selected and measured according to the procedure described in
Abbreviation, name, definition and magnification of measurements (after
|
Abbreviation |
Character name |
Definition |
Magnification in pixel/mm |
Applied to |
|
ant.l |
Antenna length |
Combined length of pedicel plus flagellum, outer aspect ( |
888 |
all species |
|
clv.b |
Clava breadth |
Greatest breadth of clava, outer aspect |
888 |
only Pc |
|
eye.b |
Eye breadth |
Greatest breadth of eye, lateral view |
888 |
only Pc |
|
eye.d |
Eye distance |
Shortest distance between eyes, dorsal view |
888 |
only Pc |
|
eye.h |
Eye height |
Greatest length of eye height, lateral view |
888 |
only Pc |
|
eye.l |
Eye length |
Length of eye, dorsal view ( |
888 |
only Pc |
|
fm3.b |
Metafemur breadth |
Greatest breadth of metafemur, outer aspect |
888 |
all species |
|
fm3.l |
Metafemur length |
Length of metafemur, from distal end of trochanter to tip of metafemur, measured along midline, outer aspect |
888 |
all species |
|
fwi.b |
Fore wing breadth |
Greatest breadth of fore wing, measured at about right angle to marginal and postmarginal veins |
444 |
only Pc |
|
fwi.l |
Fore wing length |
Greatest length of fore wing, measured from end of humeral plate to tip of wing |
444 |
only Pc |
|
gst.b |
Gaster breadth |
Greatest breadth of gaster, distance between the outermost lateral edges of the gaster, dorsal view |
444 |
all species |
|
gst.l |
Gaster length |
Length of gaster along median line from anterior edge of basal tergum to tip of ovipositor sheath, dorsal view |
444 |
all species |
|
hea.b |
Head breadth |
Greatest breadth of head, dorsal view |
888 |
all species |
|
hea.h |
Head height |
Distance between anterior margin of clypeus and anterior edge of anterior ocellus, frontal view |
888 |
all species |
|
hea.l |
Head length |
Length of head, dorsal view ( |
888 |
only Pc |
|
mav.l |
Marginal vein length |
Length of marginal vein, distance between the point at which submarginal vein touches the leading edge of the wing and the point at which the stigma vein and postmarginal vein unite ( |
888 |
all species |
|
msc.b |
Mesoscutum breadth |
Greatest breadth of mesoscutum just in front of level of tegula, dorsal view |
888 |
all species |
|
msc.l |
Mesoscutum length |
Length of mesoscutum along median line from posterior edge of pronotum to posterior edge of mesoscutum, dorsal view |
888 |
only Pc |
|
msp.l |
Malar space |
Distance between the point where malar sulcus enters mouth margin and malar sulcus enters lower edge of eye, lateral view ( |
888 |
only Pc |
|
mss.l |
Mesosoma length |
Length of mesosoma along median line from anterior edge of pronotum collar to posterior edge of nucha, dorsal view |
888 |
all species |
|
ool.l |
OOL |
Shortest distance between posterior ocellus and eye margin, dorsal view ( |
888 |
all species |
|
pdl.b |
Pedicel breadth |
Greatest breadth of pedicel, outer aspect |
888 |
only Pc |
|
pdl.l |
Pedicel length |
Length of pedicel, outer aspect |
888 |
only Pc |
|
plc.d |
Plica distance |
Greatest distance between upper edge of anterior plica ( |
888 |
all species |
|
pol.l |
POL |
Shortest distance between posterior ocelli, dorsal view ( |
888 |
all species |
|
ppd.l |
Propodeum length |
Length of propodeum measured along median line from anterior edge to posterior edge of nucha, dorsal view |
888 |
all species |
|
scp.l |
Scape length |
Length of scape exclusive of radicle, outer aspect ( |
888 |
only Pc |
|
sct.l |
Scutellum length |
Length of scutellum along median line from posterior edge of mesoscutum to posterior edge of scutellum, dorsal view |
888 |
only Pc |
|
stv.l |
Stigmal vein length |
Length of stigmal vein, distance between the point at which stigma vein and postmarginal vein unite apically and the distal end of the stigma ( |
888 |
all species |
|
tmp.l |
Temple length |
Length of temple, dorsal view ( |
888 |
only Pc |
|
upf.l |
Upper face length |
Distance between lower edge of toruli and lower edge of anterior ocellus ( |
888 |
only Pc |
We also used the Keyence microscope for making stack-images of qualitative character states. A 3-digit individual code including the notion “Palbi” (e.g. “Palbi 536”) was provided for specimens that were measured, photographed or used as reference specimens for comparison with the newly described species. Specimens already possessing a unique label from former projects (like e.g. “alb”, “Baur” etc.) or that can otherwise be recognised without a doubt (such as holotypes), were not given an additional label.
Morphometric data
To further test the hypothesis that P. capito Baur sp. n. is indeed a separate species, a shape principal component analysis (shape PCA) and a PCA ratio spectrum were applied incorporating 28 of the measurements listed above. The analysis of ratios of body measurements is an important part of morphometric taxonomy in Chalcidoidea, since many cryptic species often differ significantly in body proportions but not in qualitative characters (
Rearing
During field trips in summer 2016 in Switzerland and Italy, additional wasp specimens were collected using sweep nets (
Data resources
Morphometric raw data files, R-scripts used for calculating the shape PCA and ratios, photographs of measured and illustrated characters and species, as well as files listing examined and prepared specimens are deposited at zenodo.org. An interactive version of the Xper3 key is available under https://pteromalus.identificationkey.org.
Taxon treatment
Pteromalus capito sp. nov.
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.143853; decimalLongitude:6.795059; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:03/08/2015; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Holotype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_621; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.143853; decimalLongitude:6.795059; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:08/06/2013; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_620; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.143445; decimalLongitude:6.794963; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:08/06/2013; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_624; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.143445; decimalLongitude:6.794963; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:08/06/2013; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_625; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.144322; decimalLongitude:6.794976; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:08/06/2013; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_622; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.144322; decimalLongitude:6.794976; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:08/06/2013; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_623; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.143652; decimalLongitude:6.795199; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:08/06/2013; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_626; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.143652; decimalLongitude:6.795199; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:08/06/2013; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_627; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.143882; decimalLongitude:6.795086; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:08/06/2013; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_628; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.143882; decimalLongitude:6.795086; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:08/06/2013; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_629; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.144121; decimalLongitude:6.794942; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:08/06/2013; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_630; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.144121; decimalLongitude:6.794942; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:08/06/2013; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_631; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.14371; decimalLongitude:6.795148; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:08/06/2013; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_632; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.14371; decimalLongitude:6.795148; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:08/06/2013; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_633; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Ticino, Faido, Garlengo; decimalLatitude:46.456363; decimalLongitude:8.796262; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:08/06/2013; habitat:Marsh area; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_634; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.142351; decimalLongitude:6.795546; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:30/07/2003; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_694; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.142351; decimalLongitude:6.795546; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:30/07/2003; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_695; basisOfRecord:PreservedSpecimen
-
scientificName: Pteromalus capito; kingdom:Animalia; phylum:Arthropoda; class:Insecta; order:Hymenoptera; family:Pteromalidae; taxonRank:species; genus:Pteromalus; specificEpithet:capito; scientificNameAuthorship:Baur; continent:Europe; country:Switzerland; locality:Wallis, Champéry, Col de Bretolet; decimalLatitude:46.142351; decimalLongitude:6.795546; georeferenceProtocol:label; samplingProtocol:sweeping; eventDate:30/07/2003; habitat:Alpine meadow; individualCount:1; sex:female; lifeStage:adult; preparations:dry mounted; recordedBy:H. Baur; identifiedBy:H.Baur; dateIdentified:2017; type:Paratype; language:de; institutionID:NMBE; institutionCode:Natural History Museum Bern (NMBE); collectionCode:NMBE_HB_696; basisOfRecord:PreservedSpecimen
Description
Female: Colour: Head and mesosoma: green to blue-green with metallic lustre; setae on head and mesosoma: whitish, inconspicuous; tegula: testaceous; setae on callus of popodeum: whitish. Scape: fuscous with basal third to half testaceous; pedicel: fuscous; flagellum: brown to black. Fore wing: hyaline; fore wing venation: testaceous; setae on fore wing: whitish except for setae on the lower surface of the costal cell and for some setae along the distal part of the marginal vein which are fuscous; hind wing: hyaline. Procoxa: green-blue; meso- and metacoxa: infuscate with metallic tinge; trochanters: infuscate, testaceous at tips; profemur: infuscate, testaceous in apical sixth; mesofemur: infuscate, testaceous in apical and basal tips; metafemur: infuscate with metallic tinge, testaceous in apical tenth; pro- and mesotibiae: testaceous, medially slightly infuscate; metatibia: infuscate, basal fifth and apical third testaceous; protarsus: testaceous with distal segments infuscate; meso- and metatarsus: testaceous; pretarsi: infuscate. Petiole: dark green with purplish tinge; gaster: dark blue-green with metallic lustre; gastral terga: one to five green to blue. Sculpture: Head in frontal view: finely reticulate with moderately high septa; clypeus: striate; area between clypeus and malar sulcus: finely reticulate. Mesoscutum: finely reticulate, meshes moderately high, areoles slightly enlarged medially in posterior part of sclerite; scutellum: reticulate, meshes about as strong and coarse as on posterior part of mesoscutum; frenum: reticulate, meshes mostly similar in size to those on scutellum; axilla: reticulate, about as strongly as on central part of scutellum; prepectus upper triangular area: weakly reticulate to almost smooth; upper mesepimeron: smooth to alutaceous; upper mesepisternum: reticulate, about as strongly as on mesoscutum; metapleuron: finely reticulate, less strongly than on mesepisternum. Procoxa: alutaceous to finely reticulate; mesocoxa: finely alutaceous; metacoxa: finely reticulate. Median area of propodeum: finely reticulate, less strongly than on mesoscutum, with longitudinal ridges; inner corner of anterior plica: with a large depression, alutaceous; nucha: finely reticulate, meshes transverse, less strong than on median area of propodeum; callus of propodeum: finely reticulate; paraspiracular sulcus: smooth with few transverse costulae. Petiole in dorsal view: smooth; gastral terga: smooth and shining, second to fifth tergum anteriorly alutaceous, sixth and syntergum entirely alutaceous. Shape and structure: Head in frontal view: subtrapezoid; gena in frontal view: slighty buccate; temple in dorsal view: obtuse; forming an angle with occiput of: 110 degrees; occipital carina: absent; torulus position with respect to lower ocular line: distinctly above; lower face in lateral view: rather flat; receding with respect to upper face: weakly; forming an angle of 35 degrees; scrobe: narrow, moderately deep; malar sulcus: superficial, but traceable; clypeus anterior margin: narrowly and shallowly emarginate, without a depression above emarginate edge; gena near mouth: terete; tentorial pit: indistinct; mouth extension: not conspicuously enlarged; mandibular formula: 3-4. Antenna: Antennal formula: 11263; scape reaching: middle of anterior ocellus; flagellum: weakly clavate; first anellus: strongly transverse; second anellus: strongly transverse; first funicular segment: slightly conical; setae on flagellum: thickly clothed with subdecumbent setae, length of setae at most half as long as breadth of flagellar segments; number of rows of longitudinal sensilla on first funicular segment: 2; on sixth: 1-2. Mesosoma in lateral view: moderately bent; propodeum in lateral view sloping with respect to dorsal plane of mesoscutum and scutellum at an angle of 40 degrees; pronotum breadth with respect to mesoscutum breadth: distinctly narrower; pronotum collar: horizontal, well defined; its length with respect to mesoscutum length: slightly more than one sixth; its anterior margin: slightly elevated ridge, sometimes medially carinate; pronotum posterior margin: finely reticulate, shiny; notauli: superficial; reaching: two thirds along mesoscutum; scutellum in lateral view: moderately convex, slightly depressed around the middle; scutellum in posterior view: convex; scutellum posterior margin projection: level of anterior margin of dorsellum; scutellum posterior margin in posterior view: narrowly emarginate in the middle; frenal line: finely indicated, especially on sides; prepectus upper triangular area: separated by an indistinct oblique carina; upper mesepimeron: strongly narrowing below, not reaching base of mesopleuron; propodeum anterior plica: strong, bent inwards posteriorly to form an elevated costula; posterior plica: present, joining anterior plica; orientation of posterior plica: strongly converging; median carina of propodeum: present, often bifurcating from base of propodeum; nucha: lunate strip, clearly delimited from median area by an edge; spiracle: oval, almost touching anterior margin of propodeum; size: moderate; callus pilosity: sparsely pilose; paraspiracular sulcus: with large and moderately deep fovea slightly offset from spiracle, bluntly reflexed at level of posterior edge of fovea. Fore wing: Fore wing apex with respect to apex of gaster when folded back: slightly exceeding; basal cell number of setae: bare; basal setal line: incomplete; with 0 setae; cubital setal line: incomplete; with 0 setae; costal cell pilosity on dorsal side: bare; costal setal line: widely interrupted medially; speculum on dorsal side: bare, extending or almost extending to stigmal vein, widely open below; wing disc: sparsely pilose; marginal setae: missing; stigma: oval to subrhombical, small; uncus: short. Femora: moderately slender; metatibia: gradually widening towards apex; metacoxa pilosity dorsal: bare. Petiole in dorsal view: conical; in ventral view: open; gaster in dorsal view: elongate, acuminate; gastral terga: convex, sometimes weakly sunken; posterior margin of first gastral tergum: entire; first gastral tergum reaching: one fifth of gaster; tip of hypopygium reaching about half of gaster; ovipositor sheath: distinctly protruding. Length and body ratios: Body length: 2.8—4.0 mm; mesoscutum breadth: 635—1002 μm. Head breadth to head height: 1.30—1.45; head breadth to length: 1.89—2.25; head breadth to mesoscutum breadth: 1.02—1.15; upper face height to head height: 0.56—0.61; POL to OOL: 1.26—1.52; eye height to breadth: 1.34—1.52; eye distance to height: 1.38—1.52; temple length to eye height: 0.21—0.42; malar space to eye height: 0.46—0.54. Antenna length to head breadth: 0.86—0.94; scape length to eye height: 0.72—0.80; pedicel length to breadth: 1.33—1.94. Mesosoma length to mesoscutum breadth: 1.28—1.50; mesoscutum breadth to length: 1.43—1.87; mesoscutum length to scutellum length: 1.05—1.28; propodeum length to scutellum length: 0.29—0.42; plica distance to propodeum length: 1.90—3.05. Fore wing length to breadth: 2.15—2.36; marginal vein to stigma vein length: 1.44—1.80. Metafemur length to breadth: 3.46—3.76. Gaster length to breadth: 1.93—2.39; gaster length to mesosoma length: 1.43—1.66.
Diagnosis
The female of P. capito Baur sp. n. keys out in










P. achillei: Clypeus emargination narrow and smooth or foveate, distal part of scape fuscous or testaceous, head posterior to eye relatively narrower, sculpture of middle part of propodeum reticulate, transversely rugulose or smooth, reticulation on frenum median area coarser than or as coarse as on scutellum median area, pronotal collar one seventh to one sixth the length of mesoscutum, not elevated anteriorly, setae on fore wing fuscous, proximal half of metatibia yellow or with fuscous infumation, metacoxa only apically with setae, paraspiracular inclination sharp or blunt, area below propodeal foramen with pit present or absent, fissure absent.
P. albipennis: Clypeus emargination narrow and foveate, distal part of scape fuscous (Fig.




P. caudiger: Clypeus emargination narrow and foveate, distal part of scape fuscous, head posterior to eye relatively narrower, sculpture of middle part of propodeum reticulate or transversely rugulose, reticulation on frenum median area coarser than on scutellum median area, pronotal collar one seventh to one sixth the length of mesoscutum, usually not elevated anteriorly, setae on fore wing fuscous, proximal half of metatibia yellow or with fuscous infumation, metacoxa only apically with setae, paraspiracular inclination sharp (Fig.
P. cingulipes: Clypeus emargination narrow and smooth or foveate, distal part of scape fuscous or testaceous, head posterior to eye relatively narrower, sculpture of middle part of propodeum reticulate or transversely rugulose, reticulation on frenum median area coarser than or as coarse as on scutellum median area, pronotal collar one seventh to one sixth the length of mesoscutum, usually not elevated anteriorly, setae on fore wing pale, proximal half of metatibia with almost black infumation, metacoxa only apically with setae, paraspiracular inclination sharp (Fig.
P. eudecipiens: Clypeus emargination narrow or wide and smooth, distal part of scape fuscous or testaceous, head posterior to eye relatively narrower, sculpture of middle part of propodeum reticulate or transversely rugulose, reticulation on frenum median area coarser than or as coarse as on scutellum median area, pronotal collar one seventh to one sixth the length of mesoscutum, not elevated anteriorly, setae on fore wing fuscous, proximal half of metatibia yellow or with fuscous infumation, metacoxa only apically with setae, paraspiracular inclination sharp (Fig.
P. intermedius: Clypeus emargination wide and smooth, distal part of scape fuscous or testaceous, head posterior to eye relatively narrower, sculpture of middle part of propodeum reticulate or transversely rugulose, reticulation on frenum median area coarser than or as coarse as on scutellum median area, pronotal collar one fifth the length of mesoscutum, mostly elevated anteriorly, setae on fore wing fuscous, proximal half of metatibia yellow or with fuscous infumation, metacoxa only apically with setae, paraspiracular inclination blunt (Fig.
P. patro: Clypeus emargination narrow and foveate, distal part of scape fuscous, head posterior to eye relatively narrower, sculpture of middle part of propodeum transversely rugulose, reticulation on frenum median area coarser than on scutellum median area, pronotal collar one seventh to one sixth the length of mesoscutum, not elevated anteriorly, setae on fore wing fuscous, proximal half of metatibia yellow or with fuscous infumation, metacoxa only apically with setae, paraspiracular inclination sharp (Fig.
P. temporalis: Clypeus emargination wide and smooth, distal part of scape testaceous, head posterior to eye relatively narrower, sculpture of middle part of propodeum reticulate or transversely rugulose, reticulation on frenum median area coarser than or as coarse as on scutellum median area, pronotal collar one fifth the length of mesoscutum, usually not elevated anteriorly, setae on fore wing fuscous, proximal half of metatibia yellow, metacoxa with a few short setae also basally, paraspiracular inclination blunt (Fig.
Etymology
The species epithet “capito” refers to the relatively massive head of the species in comparison to its morphologically closest relatives. The word derives from Latin “capito” meaning “one that has a large head, big-headed”. It is a noun that does not change with gender.
Biology
Pteromalus capito Baur sp. n. is a parasitoid of larvae of Tephritidae (Diptera). Reared specimens emerged from Crepis pyrenaica (L.) Moench (coll. 07.07.2016) together with Tephritis crepidis Hendel, from Crepis paludosa (L.) Moench (coll. 21.07.2016) together with Tephritis conura (Loew), Campiglossa cf. guttella (Rond.) and Campiglossa cf. producta (Loew) and from Centaurea jacea L. s.l. (coll. 27.07.2016) together with Chaetoriella jaceae (Rob.-Desv.) and Acanthiophilus helianthi (Rossi) (Suppl. materials
Morphometric data
Fig.
Best ratios found by the LDA ratio extractor for delimiting Pteromalus capito Baur sp. n. from the most similar species, P. albipennis (alb) and P. cingulipes (cin).
|
Species comparison |
Best ratio |
Value species 1 |
Value species 2 |
|---|---|---|---|
|
capito – alb + cin |
eye.d : pol.l |
>2.73 |
<2.73 |
|
capito – alb |
ant.l : ool.l |
<5.85 |
>5.85 |
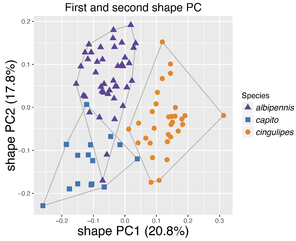
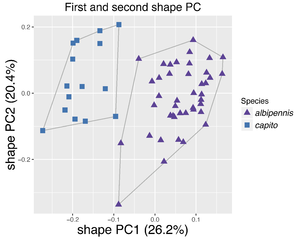
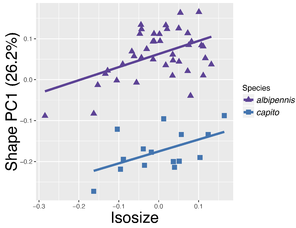
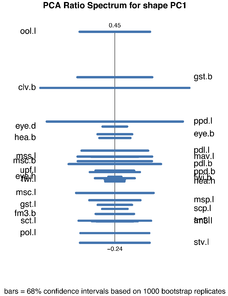
Shape PCA of females of the Pteromalus albipennis species group.
b: Scatterplot of the first against second shape PC for Pteromalus albipennis and P. capito.
c: Same species as b, but isosize plotted against first shape PC.
d: PCA ratio spectrum for the first shape PC as calculated in B and C.
The measurements for all species are listed in Suppl. materials
Analysis
New morphological characters
No study so far has examined the area of the metasternum in Pteromalidae for distinctive characters although it seems quite promising. In Fig.
The preparation of translucent slide material of the back of the head yielded no distinguishing characters. Fig.
Discussion
Interactive identification key
The development of software to analyse large amounts of descriptive data allowed the creation of new taxonomic keys, which led to a facilitated identification process. As an example, the identification of P. albipennis needs 16 steps in
It must be pointed out that the long term storage of the descriptive data and the pictures as well as the future access to the original database is a major concern. We simply do not have the experience of storing digital data over longer time periods (>20 years) (
Cryptic diversity in the P. albipennis group
It is surprising that, even in one of the taxonomically best studied areas like Central Europe, new species can be found. Pteromalus capito Baur sp. n. clearly belongs to the P. albipennis species group showing all its distinctive characters (see above). In the recent past, some new species have been described for this species group (
New morphological characters
The paraspiracular inclination has been shown to be very consistent within species and has therefore proved to be a valuable means for distinguishing especially P. capito Baur sp.n. and P. albipennis but also other species. It is not absolutely necessary to grind the specimens with a microtome when studying this character. With some experience and a good light microscope, it is possible to spot the differences without preparation. Grinding specimens was merely used to take good pictures illustrating the differences, which was otherwise impossible with the available equipment. Thirteen species showed a sharp edge of the paraspiracular inclination, of which all belong to the P. albipennis species group, in comparison to 30 species with a blunt edge (not examined for two species). As far as we know, there have been no comments in the scientific literature mentioning this structure in Pteromalidae. Studying of the area of the metasternum astonishingly revealed a whole bunch of distinctive characters. The wrinkles connecting the propodeal with the metacoxal foramen may show some sort of stabilising function of the metasternum. We assume that the impression below the propodeal foramen is linked to gaster size or ovipositor length, since species with a large gaster tend to have such an impression more often. The presence or absence of a fissure vertically below the propodeal foramen may be linked to developmental processes. The shape of the propodeal foramen, although consistent in most species, may only be an artefact of a small sample size, since we did not have the possibility to dissect a large quantity of specimens for each species. Therefore, this character may need further investigation. The metasternum has also attracted the interest of other researchers in the field of Pteromalidae.
Acknowledgements
I would like to thank Hannes Baur for his support and supervision of this work, Prof. Dr. Christian Kropf for the opportunity to write my Master thesis in his group and his kind support with the microtome, the Burgergemeinde Bern for their generous financial support of the fieldwork, Prof. Dr. Wolfgang Nentwig for a recommendation letter, the museums of Lund, Oxford and Stockholm for the loan of type specimens, Elsa Obrecht for identifying the Tephritidae species and proofreading my manuscript and, last but not least, Sina Maletti for help in collecting specimens and having a good time during the field trips. This research received support from the SYNTHESYS Project (http://www.synthesys.info) which is financed by European Community Research Infrastructure Action under the FP7 "Capacities" Programme.
References
- Can Townes type Malaise traps be improved? Some recent developments.Entomologische Berichten, Amsterdam69(4):129‑135.
- A fresh look at the reliability of long-term digital storage.ACM SIGOPS Operating Systems Review40(4):221‑234. https://doi.org/10.1145/1218063.1217957
- Baur H (2000) Monophyly and relationship of the genus Coelopisthia Foerster (Chalcidoidea, Pteromalidae). In: Austin A, Dowton M (Eds) Hymenoptera: Evolution, Biodiversity and Biological Control.CSIRO Publishing,Collingwood,165-177pp.
- Analysis of ratios in multivariate morphometry.Systematic Biology60(6):813‑825. https://doi.org/10.1093/sysbio/syr061
- Morphometric analysis and taxonomic revision of Anisopteromalus Ruschka (Hymenoptera: Chalcidoidea: Pteromalidae) – an integrative approach.Systematic Entomology39(4):691‑709. https://doi.org/10.1111/syen.12081
- Pushing the limits – two new species of Pteromalus (Hymenoptera, Chalcidoidea, Pteromalidae) from Central Europe with remarkable morphology.ZooKeys514:43‑72. https://doi.org/10.3897/zookeys.514.9910
- Fluctuating asymmetry in metric traits; a practical example of calculating asymmetry, measurement error, and repeatability.Annales Zoologici Fennici45:32‑38. https://doi.org/10.5735/086.045.0103
- Statistical methods for assessing agreement between two methods of clinical measurement.The lancet327(8476):307‑310. https://doi.org/10.1016/S0140-6736(86)90837-8
- Illustrated key to West-Palearctic genera of Pteromalidae (Hymenoptera: Chalcidoidea).Institut National de la Recherche Agronomique (INRA),Paris,140pp.
- DELTA – DEscription Language for TAxonomy.WWW documentURL: http://www.delta-intkey.com/
- Liriomyza Parasitoids in Southeast Asia. http://keys.lucidcentral.org/keys/v3/Liriomyza/
- A revision of the Encarsia pergandiella species complex (Hymenoptera: Aphelinidae) shows cryptic diversity in parasitoids of whitefly pests.Systematic Entomology42(1):31‑59. https://doi.org/10.1111/syen.12187
- Annotated Keys to the Genera of Nearctic Chalcidoidea (Hymenoptera).NRC Research Press.
- The presence of Notanisus Walker (Hymenoptera: Pteromalidae) in North America and revision of the oulmesiensis species group.Zootaxa3948(3):422‑422. https://doi.org/10.11646/zootaxa.3948.3.4
- Dutch Chalcids VIII.Entomologische. Berichten32:56‑58.
- Four new species of Pteromalus Swederus (Hymenoptera: Chalcidoidea: Pteromalidae) and redescriptions of three other species.Zoologische Mededeelingen73:165‑175.
- The Pteromalidae of North-Western Europe.Bulletin of the British Museum (Natural History), Entomology, Supplement16:1‑908.
- A new species of Pteromalus (Hymenoptera: Chalcidoidea) from France, associated with Solidago virgaurea.Entomologische Berichten51:153‑155.
- Toryminae (Hymenoptera: Chalcidoidea: Torymidae).Associated Publishers
- Janzon LÅ (1984) Taxonomy of a new subgroup of the Pteromalus albipennis group and their host relationships, with description of four new species (Hymenoptera: Chalcidoidea). Paper VI. Taxonomical and biological studies of Tephritis species (Diptera) and their parasitoids (Hymenoptera).University of Stockholm, Stockholm
- Doddifoenus wallacei, a new giant parasitoid wasp of the subfamily Leptofoeninae (Chalcidoidea: Pteromalidae), with a description of its mesosomal skeletal anatomy and a molecular characterization.Zootaxa2194:21‑36.
- Multivariate ratio analysis reveals Trigonoderus pedicellaris Thomson (Hymenoptera, Chalcidoidea, Pteromalidae) as a valid species.Systematic Entomology38:753‑762. https://doi.org/10.1111/syen.12026
- Phylogenetic analysis of Eurytominae (Chalcidoidea: Eurytomidae) based on morphological characters.Zoological Journal of the Linnean Society151(3):441‑510. https://doi.org/10.1111/j.1096-3642.2007.00308.x
- Taxonomic differentiation of three species of the Pteromalus albipennis species group (Hymenoptera: Chalcidoidea: Pteromalidae), using an integrative approach.Bachelor thesis, Rheinische-Fridrich-Wilhelms Universität, Bonn1‑32.
- From morphometry to taxonomy – using multivariate analyses for the evaluation and characterization of cryptic species.Insect Systematics & Evolution.
- An interactive key to the Heterospilus (Hymenoptera: Braconidae: Doryctinae) of Costa Rica. http://keyserver.lucidcentral.org/key-server/player.jsp?keyId=106
- Collecting and preserving chalcid wasps (Hymenoptera: Chalcidoidea).Journal of Natural History16(3):315‑334. https://doi.org/10.1080/00222938200770261
- Universal Chalcidoidea Database. World Wide Web electronic publication. http://www.nhm.ac.uk/chalcidoids
- R: A language and environment for statistical computing.R Foundation for Statistical Computing, Vienna, Austria. URL: http://www.R-project.org
- NIH Image to ImageJ: 25 years of image anaylsis.Nature Methods9(7):671‑675. https://doi.org/10.1038/nmeth.2089
- Desktop or remote knowledge base management systems for taxonomic data and identification keys: Xper2 and Xper3.Proceedings of TDWG1:e19911. https://doi.org/10.3897/tdwgproceedings.1.19911
- ggplot2: Elegant Graphics for Data Analysis.Springer-Verlag, New York.
- Immunohistochemical localization of cell surface receptors using a novel method permitting simple, rapid and reliable LM/EM correlation.The Histochemical Journal18(7):387‑396. https://doi.org/10.1007/BF01675220
Supplementary materials
All labelled and measured specimens relevant for this work are listed with their sample date, location, deposition, manipulations etc.
Body measurements for every single individual used for the key. This file includes 15 measurements and 8 ratios.
This file contains the Min and Max values for each species included in the key for the body measurements.
This file contains the Min and Max values for every species in the key for each of the 8 calculated ratios.
This files lists all specimens that were treated with the microtome for the examination of the paraspiracular inclination including information of taxon, deposition, sex, sampling data, sampling location etc.
This file lists all flower head samples that have been collected during summer 2016 and from which Chalcidoids were reared, including information on sampling date, sampling location, plant species, number and taxon of insects hatched etc.
This file lists all species belonging to Chalcidoidea or Tephritidae which hatched from the collected flower heads.
This file contains all measurements (also from former gaugers) used by the authors for the Bland-Altman training, which was performed prior to measuring specimens relevant to the key.
This files lists all body measurements, ratios and their Min and Max values for each specimen of P. capito Baur sp. n. used for the Principal Component Analysis and the key.
This file contains all relevant data for the Principal Component Analysis and for the creation of the plots used in this paper.
This R file contains the code for the Principal Component Analysis and the creation of the plots used in this work.