|
Biodiversity Data Journal :
Taxonomic paper
|
Filling in the gap: two new records and an updated distribution map for the Gulf Sand gecko Pseudoceramodactylus khobarensis Haas, 1957
|
Corresponding author:
Academic editor: Truong Nguyen
Received: 02 Sep 2014 | Accepted: 01 Oct 2014 | Published: 06 Oct 2014
© 2014 Margarita Metallinou, Raquel Vasconcelos, Jiří Šmíd, Roberto Sindaco, Salvador Carranza
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Metallinou M, Vasconcelos R, Šmíd J, Sindaco R, Carranza S (2014) Filling in the gap: two new records and an updated distribution map for the Gulf Sand gecko Pseudoceramodactylus khobarensis Haas, 1957. Biodiversity Data Journal 2: e4011. https://doi.org/10.3897/BDJ.2.e4011
|
|
Keywords
Reptilia, Gekkonidae, DNA, 12S, distribution range, Arabia, sabkha
Introduction
The genus Pseudoceramodactylus Haas, 1957 comprises a single species, the Gulf Sand gecko P. khobarensis, described from eastern Saudi Arabia (



Specimen and habitat images of Pseudoceramodactylus khobarensis from western central Oman.
b: Detail of the left side of the head of the same specimen, where it is possible to observe the swollen nasal area.
c: Interdune sabkha in the general area where a juvenile P. khobarensis was collected, north of Hasirah oil field, in central-western Oman. Both specimens were found active during the night (Table
Pseudoceramodactylus khobarensis was transferred to the genus Stenodactylus by
Herein, we report two new records for this species from the eastern edge of the Rub Al Khali desert (‘Uruq al Mu’taridah area), in inner Oman (Fig.
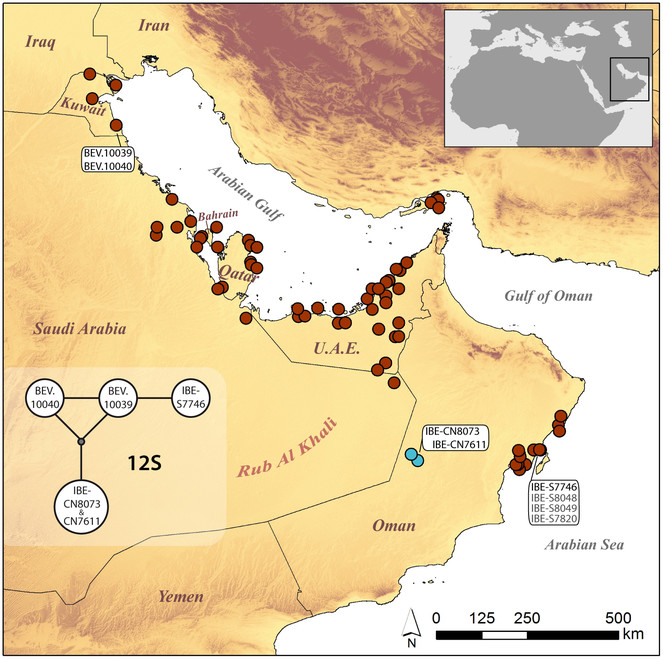
Updated distribution map for Pseudoceramodactylus khobarensis, with new records in light blue color. Localities of material examined in this study are indicated and corresponding specimen codes are provided. In the inset figure, the haplotype network of the mitochondrial marker 12S is presented. Exact localities, 12S GenBank accession codes and morphological measurements of specimens are listed in Table
New records of localities where P. khobarensis was collected in eastern Rub Ak Khali desert, Oman.
| Specimen Code | Date | Time | Latitude | Longitude | Elevation (m) | Temperature (°C) | Humidity | Other species collected |
|---|---|---|---|---|---|---|---|---|
| IBE-CN8073 | 7-Oct-2013 | 02:00 am | 20.66029 | 55.54131 | 96 | 27.8 | 50.2 | S. arabicus |
| IBE-CN7611 | 7-Oct-2013 | 00:30 am | 20.50214 | 55.69896 | 143 | 28.3 | 57.6 | none |
Morphometrical (in mm) and meristic measurements for the specimens examined in this study, originating from four different localities across the range of P. khobarensis (see Fig.
| Specimen Code | IBE-CN8073 | IBE-CN7611 | IBE-S7746 | IBE-S8048 | IBE-S8049 | IBE-S7820 | BEV.10039 | BEV.10040 |
|---|---|---|---|---|---|---|---|---|
| GenBank 12S | KM047415 | KM047415 | KC190704 | - | - | - | KC190703 | KC190702 |
| Latitude | 20.6603 | 20.5021 | 20.6854 | 20.6854 | 20.6854 | 20.6854 | 28.6369 | 28.6369 |
| Longitude | 55.5413 | 55.699 | 58.2934 | 58.2934 | 58.2934 | 58.2934 | 48.1336 | 48.1336 |
| Sex | (juvenile) | male | male | male | male | female | male | female |
| Snout-vent length (SVL) | 28.52 | 61.37 | 50.90 | 53.56 | 53.05 | 50.72 | 50.71 | 56.80 |
| Head length (HL) | 8.28 | 15.08 | 13.17 | 14.34 | 14.69 | 13.00 | 13.29 | 14.50 |
| Head width (HW) | 4.78 | 7.93 | 6.17 | 6.56 | 6.93 | 6.21 | 7.25 | 7.80 |
| Eye diameter (ED) | 2.60 | 4.28 | 4.02 | 3.74 | 4.07 | 4.15 | 3.63 | 4.17 |
| Forearm length (FL) | 7.72 | 12.61 | 11.36 | 10.96 | 11.48 | 11.64 | 11.37 | 11.59 |
| Arm length (AL) | 4.54 | 9.19 | 7.32 | 7.62 | 8.24 | 7.78 | 7.76 | 8.10 |
| Tibia length (BL) | 7.64 | 13.79 | 12.39 | 11.91 | 13.43 | 13.05 | 11.55 | 13.68 |
| Femur length (ML) | 6.66 | 13.63 | 10.50 | 11.54 | 13.32 | 11.86 | 10.61 | 12.10 |
| Tail length (TL) | 27.13 | 53.74 | 36.19 | 43.56 | 40.04 | 38.76 | N/A | 47.45 |
| Upper labials (ULS) | 13 | 13 | 12 | 12 | 11 | 12 | 12 | 12 |
| Lower labials (LLS) | 10 | 12 | 10 | 11 | 10 | 11 | 12 | 11 |
Materials and methods
During intensive fieldwork in Oman, in October 2013, we surveyed the easternmost tip of the Rub Al Khali desert (Fig.
Data for the updated distribution map were compiled from
A total of five individuals, the two newly collected ones and three additional specimens from the extremes of the species’ range (Fig.
A series of morphological measurements were performed on the same five individuals, as well as three additional specimens from the locality in coastal Oman belonging to the field series of S. Carranza (Table
Taxon treatment
Pseudoceramodactylus khobarensis
- Reptile Database http://reptile-database.reptarium.cz/species?genus=Pseudoceramodactylus&species=khobarensis
- Encyclopedia of Life http://eol.org/pages/461035/overview
- ITIS http://www.itis.gov/servlet/SingleRpt/SingleRpt?search_topic=TSN&search_value=819426
-
taxonID: ; scientificNameID: urn:lsid:organismnames.com:name:2791139; country:Oman; stateProvince:Al Wusta; verbatimLocality:north of Hasirah oil field, ‘Uruq al Mu’taridah area; verbatimElevation:143 m; verbatimLatitude:20 30 7.704N; verbatimLongitude:55 41 56.2554E; eventDate:2013-10-07T00:30+0400; individualCount:1; sex:male; recordedBy:Salvador Carranza; Raquel Vasconcelos; Margarita Metallinou; Roberto Sindaco; Jiri Smid; collectionID:IBE-CN7611; institutionCode:Institute of Evolutionary Biology (CSIC - Universitat Pompeu Fabra)
-
taxonID: ; scientificNameID: urn:lsid:organismnames.com:name:2791139; country:Oman; stateProvince:Al Wusta; verbatimLocality:about 13km by air east of Sahmah oil filed, ‘Uruq al Mu’taridah area; verbatimElevation:96 m; verbatimLatitude:20 39 37.0434N; verbatimLongitude:55 32 28.716E; eventDate:2013-10-07T02:00+0400; individualCount:1; sex:juvenile; recordedBy:Salvador Carranza; Roberto Sindaco; Margarita Metallinou; Raquel Vasconcelos; Jiri Smid; collectionID:IBE-CN8073; institutionCode:Institute of Evolutionary Biology (CSIC - Universitat Pompeu Fabra)
Analysis
Analysis of the mitochondrial 12S marker revealed that both newly collected specimens share the same haplotype. Along a 380-bp alignment, there are 3 differences compared to sample IBE-S7746 from Barr Al-Hickman, in coastal Oman (M196 in
The mean SVL of the specimens measured was 53.87 mm (50.71–61.37, N=7), and did not differ for males (53.92, 51.71–61.37, N=5) and females (53.76, 50.72–56.80, N=2). Tail length measured between 71.1 and 95.1% of SVL.
Discussion
The Rub Al Khali is the largest desert in Arabia, the largest sand desert in the world and one of the driest (
Interestingly, the low variability of the mtDNA observed with the sequenced marker (12S) indicates that there is probably connectivity between populations across its distribution range and corroborates the hypothesis that this species inhabits larger inland areas. Indeed, coastal and inland sabkhas are abundant in eastern Saudi Arabia (
Based on the measurements performed on voucher specimens in this study, Pseudoceramodactylus khobarensis is shown to have substantially higher maximum SVL than previously documented. One female reached 56.80 mm (BEV.10040) and one male 61.37 mm (IBE-CN7611), both exceeding SVL of the largest specimens measured by
Pseudoceramodactylus khobarensis is a remarkable desert reptile in that it is the only lizard habitually found on sabkha substrate (
Acknowledgements
The authors would like to thank Erika Zakar for laboratory assistance and Philippe Geniez and Pierre-André Crochet for facilitating access to the BEV collection of the team Biogéographie et Écologie des Vertébrés de l’École Pratique des Hautes Études, Centre d'Écologie Fonctionelle et Evolutive, Montpellier, France. Special thanks are also due to Saleh Al Saadi, Mohammed Al Shariani, Thuraya Al Sariri, Ali Al Kiyumi, Mohammed Abdullah Al Maharmi and the other members of the Nature Conservation Department of the Ministry of Environment and Climate, Sultanate of Oman for their help and support and for issuing collecting permits for this survey (Ref: 21/2013). This work was supported by the project “Field study for the conservation of reptiles in Oman” funded by the Ministry of Environment and Climate Affairs, Oman (Ref: 22412027), and grant CGL2012-36970 from the Ministerio de Economía y Competitividad, Spain (co-funded by FEDER). M.M. was supported by a FPU predoctoral grant from the Ministerio de Educación, Cultura y Deporte, Spain (AP2008-01844). R.V. is supported by a postdoctoral grant from the Fundação para a Ciência e Tecnologia (FCT) (SFRH/BPD/79913/2011). The work of J.S. was supported by the Ministry of Culture of the Czech Republic (DKRVO 2014/14, National Museum, 00023272) and by the SVV 260 087/2014 project of the Charles University in Prague.
References
- Little-known geckoes (Reptilia: Gekkonidae) from Arabia with descriptions of two new species from the Sultanate of Oman.The Scientific Results of the Oman Flora and Fauna Survey 197581‑110.
- Reptiles of Saudi Arabia: A Review of the Lizard Genus Stenodactylus (Reptilia: Gekkonidae).Fauna of Saudia Arabia2:368‑404.
- Median-joining networks for inferring intraspecific phylogenies.Molecular biology and evolution16(1):37‑48. https://doi.org/10.1093/oxfordjournals.molbev.a026036
- Barth H (2002) The sabkhat of Saudi Arabia - An Introduction. In: Barth H, Böer B (Eds) Sabkha Ecosystems: Volume I: The Arabian Peninsula and Adjacent Countries.36.Springer,368pp.
- Notebook. Reptile Records.Bulletin of the Emirates Natural History Group10.1:22‑23.
- Stenodactylus khobarensis (Haas, 1957): An addition to the Iranian herpetofauna (Reptilia: Squamata: Gekkonidae).Russian Journal of Herpetology14(3):229‑231.
- Arabian Deserts: Nature, Origin and Evolution.Springer,Dordrecht, The Netherlands,592pp. https://doi.org/10.1007/1-4020-3970-0
- Arcmap 10.Environmental Systems Research InstituteRedlands:CA.
- Molecular systematics of Stenodactylus (Gekkonidae), an Afro-Arabian gecko species complex.Molecular phylogenetics and evolution58(1):71‑75. https://doi.org/10.1016/j.ympev.2010.10.014
- Gallagher MD (1971) The Amphibians and Reptiles of Bahrain. British Museum (Natural history).40pp.
- The Amphibians and Reptiles of Oman and the UAE.Chimaira,Frankfurt am Main,480pp. [ISBN978-3-89973-433-1]
- From Rifted Margins to Foreland Basins: Investigating Provenance and Sediment Dispersal Across Desert Arabia (Oman, U.A.E.).Journal of Sedimentary Research73(4):572‑588. https://doi.org/10.1306/101702730572
- Some amphibians and reptiles from Arabia.Proceedings of the California Academy of Sciences29(3):47‑86.
- Recent developments in the MAFFT multiple sequence alignment program.Briefings in bioinformatics9(4):286‑298. https://doi.org/10.1093/bib/bbn013
- Higher taxonomic categories of gekkonid lizards and their evolution.Bulletin of the American Museum of Natural History135:1‑60.
- Plant life in the Umm as Samim, Oman – A case study in a major inland sabkha.Journal of Arid Environments85:122‑127. https://doi.org/10.1016/j.jaridenv.2012.06.007
- Martens H (1996) A preliminary survey of the terrestrial reptile and sea snakes in the Jubail Marine Wildlife Sanctuary. In: Krupp F, Abuzinada AH, Nader IA (Eds) A marine wildlife sanctuary for the Arabian Gulf: environmental research and conservation following the 1991 Gulf War Oil Spill.NCWCD and Senckenberg Research Institute,Riyadh and Frankfurt.
- Herpetological observations within the framework of national park planning: Barr al Hikman and Masirah Island, Sultanate of Oman (Amphibia, Reptilia).Faunistische Abhandlungen (Dresden)21:89‑97.
- New species of Stenodactylus (Squamata: Gekkonidae) from the Sharqiyah Sands in northeastern Oman.Zootaxa3745(4):449‑468.
- Conquering the Sahara and Arabian deserts: Systematics and biogeography of Stenodactylus geckos (Reptilia: Gekkonidae).BMC Evolutionary Biology12(258):1. https://doi.org/10.1186/1471-2148-12-258
- Reptiles.Tribulus4(1):29.
- Pseudoceramodactylus khobarensis. In: IUCN 2014. IUCN Red List of Threatened Species.2014.1. Release date:2014-6-25. URL: http://www.iucnredlist.org/details/164699/0
- The reptiles of the Western Palearctic.Edizioni Belvedere,Latina (Italy),579pp. [ISBN978-88-89504-14-7]
- MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods.Molecular biology and evolution28(10):2731‑2739. https://doi.org/10.1093/molbev/msr121
- On the presence and distribution of the Gulf sand gecko, Pseudoceramodactylus khobarensis Haas, 1957 (Reptilia: Squamata: Gekkonidae) in Qatar.QScience Connect2013(34):1.
- Saudi Arabia: An environmental review.CRC Press,Leiden,332pp. [ISBN9780415413879] https://doi.org/10.1201/9780203030882