|
Biodiversity Data Journal :
Taxonomic paper
|
The Afrotropical Miomantis caffra Saussure 1871 and M. paykullii Stal 1871: first records of alien mantid species in Portugal and Europe, with an updated checklist of Mantodea in Portugal (Insecta: Mantodea)
|
Corresponding author:
Academic editor: Edward Baker
Received: 28 Sep 2014 | Accepted: 30 Oct 2014 | Published: 12 Nov 2014
© 2014 Eduardo Marabuto
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Marabuto E (2014) The Afrotropical Miomantis caffra Saussure 1871 and M. paykullii Stal 1871: first records of alien mantid species in Portugal and Europe, with an updated checklist of Mantodea in Portugal (Insecta: Mantodea). Biodiversity Data Journal 2: e4117. https://doi.org/10.3897/BDJ.2.e4117
|
|
Abstract
The recent growing interest on the Mantodea fauna of southern Europe and Portugal in particular, has enabled the discovery of two geographically separated populations of hitherto unknown species in Europe. Analysis of specimens shows that they belong to two Afrotropical mantids: Miomantis caffra Saussure, 1871 and Miomantis paykullii Stal, 1871, thus raising the number of known species in Europe to 39 and in Portugal to 11.
While these are remarkable findings, they also represent the first alien mantis species recorded from this continent. As yet, these species appear to be confined to artificial humanised gardened areas but call for more attention to the problem of biological invasions and the need for better bio-security measures for the conservation of natural ecosystems.
In the absence of recent revisionary work on the Mantodea of Portugal and given the need to provide an accessible identification tool, both a checklist and a key to species are provided for all species in the country.
Keywords
New records, Western Europe, Biological invasion, biotic homogenisation, mantis, key to species
Introduction
Human impact on biological diversity can occur at different levels and magnitudes. The effect on ecosystems can be direct, by their removal or alteration, or manifest themselves indirectly such as through climate change or the translocation of species between different biogeographic regions. Generally, due to an incomplete knowledge of ecosystems by researchers and policy-makers, many introduction cases may be overlooked, even more rarely tracked down precisely as they happen. Therefore, their effect has mostly been under-evaluated. Competition with natives for resources and other environmental constraints limits the successful establishment of most alien species (
The term "alien species" generally includes those naturally exogenous to a given habitat, ecosystem or biogeographical area which have established themselves outside their natural range either unaided or with human assistance. Several paths and vectors for introduction of such species have been identified (e.g.
In Europe there are over 1500 established alien arthropod species, of which 1390 are insects (
While mantises have so far not been reported as alien species in Europe, the worldwide scenario is quite different. From as early as 1899, Mantis religiosa L. was introduced into USA and Canada with nursery plants (
The European Mantodea species list (
Irrespective of the criteria for the definition of species and biogeographical structure, the most up-to-date and synthetic overall work on Euro-Mediterranean Mantodea,
In the summer of 2014, unexpected findings of two mantis species in Portugal triggered a revision of the species occurring in the country, alongside the investigation of their identity. In fact, the specimens studied do not key to any of the known European species of Mantodea. A combination of characters such as complete development of wings, simple cerci, no foliaceous leg extensions, small size and an elongated pronotum (in contrast to all the Ameles spp.) among others place the available specimens in the genus Miomantis.
Miomantis Saussure, 1870 currently includes 70 Afrotropical species (
Materials and methods
The first specimen of M. paykullii was collected by hand at a garden light around which it was flying. A second specimen was observed 3 days later in the same place by beating the vegetation (Arbutus unedo and Phyllostachys aurea bamboo hedge) but flew off to an inaccessible area. The third and fifth to seventh specimens, belonging to M. caffra were attracted to a porch light during the night and intercepted hunting insects attracted by it. The fourth specimen, also M. caffra, was located in a different locality. All six collected specimens of Miomantis spp. were maintained alive till further analysis. Later, these were prepared and mounted dry, remaining in the author's personal reference collection. The captured specimen of M. paykullii and the first of M. caffra were photographed by the author.
Taxon treatments
Miomantis paykullii
-
taxonRank: species; vernacularName:Egyptian pygmy mantis; genus:Miomantis; specificEpithet:paykullii; scientificNameAuthorship:Stal, 1871; continent:Europe; country:Portugal; countryCode:PT; stateProvince:Faro; county:Loulé; municipality:Quarteira; locality:Vila Sol; verbatimElevation:40; decimalLatitude:37.090; decimalLongitude:-08.093; samplingProtocol:ad hoc observation; eventDate:2014-08-05; habitat:garden; individualCount:1; sex:male; recordedBy:Eduardo Marabuto
-
taxonRank: species; vernacularName:Egyptian pygmy mantis; genus:Miomantis; specificEpithet:paykullii; scientificNameAuthorship:Stal, 1871; continent:Europe; country:Portugal; countryCode:PT; stateProvince:Faro; county:Loulé; municipality:Quarteira; locality:Vila Sol; verbatimElevation:40; decimalLatitude:37.090; decimalLongitude:-08.093; samplingProtocol:vegetation beating; eventDate:2014-08-08; habitat:garden; individualCount:1; sex:male; recordedBy:Eduardo Marabuto
Description
Adapted from
Both observed specimens are adult-stage males conforming well with the descriptions of the species available (




Miomantis paykullii, observed and photographed in Algarve, Portugal.
b: Dorsal view. Note the smooth lateral side of relatively long pronotum, straight frons, simple cerci and hyaline wings.
c: Head frontal view. Note the conical, prominent and striped eyes.
d: Detail of foreleg. Characteristic are the coxa bearing very small spines (5-6), femora crenulate between the 4 external spines and tibiae with 7 external spines.
Distribution
An Afrotropical species cited throughout the biogeographical area, with some island populations. Countries where it has been found are: Burkina Faso, Cameroon, Chad, Egypt, Ghana, Israel, Ivory Coast, Kenya, Mauritania, Mauritius, Mozambique, Niger, Senegal, Togo, Uganda and Zimbabwe (
The area is a private property garden of the resort Vila Sol where many exotic plant species are planted. Among them, hedges of Phyllostachys aurea and Lantana camara are typical while stands of Pennisetum alopecuroides, Cyperus papyrus and garden turf are widespread with smaller amounts of other exotic species. The only natives are isolated Pinus pinea trees, remnants from the open woodland pre-resort and a hedge of Arbutus unedo. At dusk, several garden lights at ground level are regularly lit and during the night, an automatic irrigation system maintains humidity levels high, even during the summer. Prior to the installment of the resort, the whole area was a dry thermomediterranean open Pinus pinea woodland with mild winters and warm summers on an arenite substrate. The biogeographic province is the Gaditano-Onubo-Algarviense in its Algarviense coastal sector with some notable endemics (
Ecology
Ecological plasticity in this species over a broad temperature range accounts for its wide distribution. According to
Conservation
Showing a wide distribution centred in the subsaharan African continent, M. paykullii should not be at risk of any kind. However, in the Euro-Mediterranean area, where it has only been found along the Nile valley and nearby areas of Israel, this species has recently been evaluated as at "Potential risk", because of sparse observations for a long time (
Miomantis caffra
-
taxonRank: species; genus:Miomantis; specificEpithet:caffra; scientificNameAuthorship:Saussure, 1871; continent:Europe; country:Portugal; countryCode:PT; stateProvince:Lisboa; county:Cascais; municipality:Carcavelos; locality:São Miguel das Encostas; verbatimElevation:50; decimalLatitude:38.701; decimalLongitude:-9.336; samplingProtocol:ad hoc observation; eventDate:2014-09-09; habitat:garden; individualCount:1; sex:male; recordedBy:Eduardo Marabuto
-
taxonRank: species; genus:Miomantis; specificEpithet:caffra; scientificNameAuthorship:Saussure, 1871; continent:Europe; country:Portugal; countryCode:PT; stateProvince:Estremadura; county:Lisboa; municipality:Oeiras; locality:Quinta do Marquês; verbatimElevation:35; decimalLatitude:38.696; decimalLongitude:-9.329; samplingProtocol:ad hoc observation; eventDate:2014-09-20; habitat:garden; individualCount:1; sex:male; recordedBy:Eduardo Marabuto
-
taxonRank: species; genus:Miomantis; specificEpithet:caffra; scientificNameAuthorship:Saussure, 1871; continent:Europe; country:Portugal; countryCode:PT; stateProvince:Lisboa; county:Cascais; municipality:Carcavelos; locality:São Miguel das Encostas; verbatimElevation:50; decimalLatitude:38.701; decimalLongitude:-9.336; samplingProtocol:ad hoc observation; eventDate:2014-09-22; habitat:garden; individualCount:2; sex:male; recordedBy:Eduardo Marabuto
-
taxonRank: species; genus:Miomantis; specificEpithet:caffra; scientificNameAuthorship:Saussure, 1871; continent:Europe; country:Portugal; countryCode:PT; stateProvince:Lisboa; county:Cascais; municipality:Carcavelos; locality:São Miguel das Encostas; verbatimElevation:50; decimalLatitude:38.701; decimalLongitude:-9.336; samplingProtocol:ad hoc observation; eventDate:2014-09-27; habitat:garden; individualCount:1; sex:male; recordedBy:Eduardo Marabuto
Description
Adapted from
The six observed and collected individuals of M. caffra are adult specimens found by porch lights at night. All are identical in pattern and therefore only the first is described, while measurements are also given for the remainder. It is a match for the description in




First specimen of Miomantis caffra, observed and photographed in Carcavelos, Portugal.
b: Dorsal view. Note the smooth lateral side of relatively long pronotum, bulging globular eyes, longer wings than abdomen extending beyond cerci.
c: Head, frontal view. Note the round, bulging eyes.
d: Detail of inner part of forelegs. Characteristic are slightly spotted coxae (5-6 black patches) and femora (3 patches).
Distribution
Originally endemic to the extreme south of Africa in South Africa and Mozambique. Original area spans from Cape of Good Hope to Maputo Bay, the former Transvaal province and Natal (
The area surrounding sightings of M. caffra is a suburban neighbourhood where managed gardens are abundant. Paspalum spp. lawns are widespread interspersed with stands of Lantana camara, Pittosporum tobira, Yucca aloifolia, Nerium oleander and cultivated Rosa spp. Other widely planted exotic species include Buxus sempervirens, Ligustrum ovalifolium, Phoenix canariensis and Hibiscus spp. Such gardens are watered every night and the whole environment contrasts with natural vegetation types. Here, the natural series would be a meso-thermomediterranean xerothermophilous vegetation adapted to a limestone substrates, the woodland series Arisaro clusii- Querco broteroi sigmetum, typical of western Portugal (
Ecology
All from
Updated checklist of the Mantodea in Portugal
Family
Perlamantis allibertii
In Portugal this species is known from a wide inland area throughout the whole country.
Family
Empusa pennata
A widespread species in Portugal, found throughout the country.
Family
Ameles spallanzania
In Portugal a regular and widespread species.
Ameles paradecolor
Throughout the whole of Portugal but uncommon, favouring Mediterranean inland areas.
Apteromantis aptera
Only locally common species, limited to the southern half of Portugal in Mesomediterranean open areas.
Geomantis larvoides
In Portugal, throughout the country but preferring sandy areas near the coast.
Mantis religiosa
Common and throughout the whole of Portugal in both natural and urban areas.
Miomantis paykullii
Extreme south of Portugal in the Algarve under subtropical semi-natural conditions. Only in the Quarteira area (Loulé, Faro, Algarve) based on two specimens.
This work
Miomantis caffra
Suburban area of Lisboa, Portugal in anthropised semi-natural conditions. Only in the area of Carcavelos and Oeiras (Cascais, Lisboa), based on five male specimens.
This work.
Sphodromantis viridis
Only known from inland areas near the border with Spain in the southern half of Portugal.
Family
Iris oratoria
Throughout Portugal but more abundant at the end of summer and in the Mediterranean areas of the south.
Identification keys
|
Key to Portuguese species of Mantodea This key is to be used with specimens in the adult stage. In nymphs, the absence of developed wings may lead to erroneous identifications. |
||
| 1 | Wings absent | 2 |
| – | Wings present | 3 |
| 2 | Eyes conical or pointed. Green to yellowish brown coloration. | Apteromantis aptera (Fuente, 1894) |
| – | Eyes globular, round shape. Body dark grey to brown, mottled and abdomen bearing a dorsal stripe. | Geomantis larvoides Pantel, 1896 |
| 3 | Pronotum short, <2x or = length of head | Perlamantis allibertii Guérin-Méneville, 1843 |
| – | Pronotum longer than 2x length of head | 4 |
| 4 | Head with a conical process, mid and hind-femora and abdomen underside with tubercles or foliose projections | Empusa pennata (Thunberg, 1815) |
| – | Head and body without such structures | 5 |
| 5 | Forewing long, reaching the tip of the abdomen, with a central white or cream-coloured spot (stigma) | Sphodromantis viridis (Forskal, 1775) |
| – | Forewing long or short, without white or cream area | 6 |
| 6 | Hindwing with a large dark coloured, blue sheen ocellus in the anal field. This is often surrounded by red, yellow or orange areas. Wings to tip of abdomen in male, shorter in female. | Iris oratoria (Linnaeus, 1758) |
| – | Hindwings without ocellus, monochromatic. If not transparent, yellow, black or red. | 7 |
| 7 | Dark spot on internal fore-coxa, often centred yellow or white and exposed during deimatic display | Mantis religiosa (Linnaeus, 1758) |
| – | Without dark spot in the fore coxa | 8 |
| 8 | Body length > 4x pronotum length. Stouter species with thicker pronotum and raptorial forelegs. Wings fully developed in male, very short in female. | 9 Genus Ameles |
| – | Body length < 4x pronotum length. Slender species with prominent eyes. Fully winged, in male reaching or passing tip of abdomen, in female shorter. | 10 |
| 9 | Ratio pronotum length / maximum width <2. Eyes ovoid with an apical spine. | Ameles spallanzania (Rossi, 1792) |
| – | Ratio pronotum length / maximum width >2. Eyes rounded and not prominent. | Ameles paradecolor Agabiti, Salvatrice & Lombardo, 2010 |
| 10 | Eyes conical and head appearing compressed dorsally. Wings hyaline in male, exceeding only slightly the tip of abdomen, but not cerci. In female, slightly shorter than abdomen. Abdomen uniform colour dorsally and ventrally. Unpatterned frontal raptorial legs. | Miomantis paykullii Stal, 1871 |
| – | Eyes globular. Wings hyaline in male exceeding tip of cerci. In female, much shorter than tip of abdomen. Abdomen dichromatic, yellow dorsally. Raptorial forelegs with small black dots especially in femora. | Miomantis caffra Saussure, 1871 |
Analysis
The first Portuguese and European specimens of wild-caught mantises in the genus Miomantis were observed in late Summer 2014.
Miomantis paykullii, was located in Vila Sol, Quarteira, Algarve, Portugal from two observations separated by three days in August. The second specimen escaped, preventing further inspection other than sexing and identification. One month later, six males of Miomantis caffra were collected in the area of Carcavelos/Oeiras, Lisboa, Portugal at fluorescent porch lights to which moths, lacewings and beetles (potential prey) are regularly attracted. Known distribution of both species in Portugal is now represented in Fig.
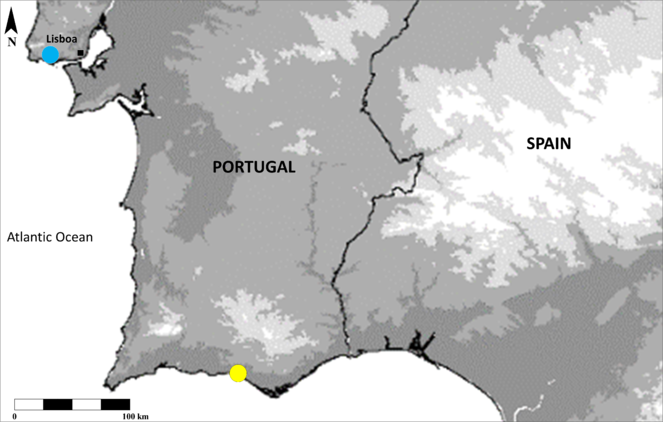
To accomodate these findings, the 11 species of Mantodea in Portugal are revised giving the general distribution of each species in the country and citing relevant literature. With the aim of supporting further studies and aiding recording, a key for the identification of adult specimens of all species within the country (view Identification key) is included.
Discussion
With the discovery of both Miomantis paykullii and Miomantis caffra in Portugal during the summer of 2014, there are now 11 mantis species known in the country and 39 in the European continent. The biogeographical realm of the genus Miomantis is the Afrotropical region, at least 2000 km away from Portugal and where all the 70 known species are endemic.
The straightforward assignment of specimens to known and relatively widespread taxa and their discovery in heavily modified habitats in European territory probably indicate faunistic novelty rather than natural relictual occurrence. M. paykullii and M. caffra are therefore, the first alien mantises in the European continent and are among the relatively few other cases known in the world (see introduction).
The Euro-Mediterranean biome is a renowned hotspot for biological diversity (
These findings enhance and are derived from the increasing pressure biodiversity is facing with rising worldwide biotic homogenisation (
All Portuguese Miomantis spp. were collected in completely man-engineered habitats buffered from Mediterranean habitats. In these gardens, commercial and widespread garden plants are used and watering is regular, establishing a subtropical environment and hampering many native species to settle, compete or interfere.
The first hypothesis to their origin includes long range natural dispersal from original distribution areas. This would involve a range expansion of from 2000 to 4000 km by M. paykullii from either Mauritania or Egypt. In this case, man-made habitats like gardens and parks would act as stepping stones overcoming biogeographical barriers. This is even less likely for M. caffra which is endemic to South Africa. Even though gardens, parks and suitable areas are fairly continuous in Europe and maybe the Maghreb, there are still wide expanses of unsuitable (either too dry or sparsely populated) ground to allow these species to spread naturally. Moreover, reaching the Iberian Peninsula as the leading edge of a widening distribution area would indicate a presence in countries in-between, which has not yet been verified.
In the second hypothesis, worldwide trade allows for a fast and intense exchange of goods, easily overcoming natural biogeographic boundaries. All developmental stages but especially oothecae attached to solid surfaces and nymphs on potted plants would be easily transportable. This has happened before, as in the case of the Japanese mantis Statilia maculata reaching New Zealand (
Beside the potential threat an alien predator such as Miomantis spp. may impose on potential prey species (i. e. all smaller arthropods), these mantises could also compete with autochthonous species. In Portugal, the only species that venture regularly into gardens and parks are Mantis religiosa and to a much lesser extent Iris oratoria and Empusa pennata. These species attain on average a larger size and are more strongly built than Miomantis spp., meaning that direct predation by the alien upon them is unlikely. However, Miomantis species are aggressive generalists which might prey on the same insects and thus out-compete natives. M. caffra itself has proven to be displacing Orthodera novaezelandiae, the only New Zealand native mantis in the outskirts of Auckland (
Whether these Miomantis populations are already established or are just the result of episodic releases or escapees, the potential disturbance to the ecosystem requires immediate monitoring and research. The extent of their influence is for now unknown as well as the means of their arrival and therefore a call for action is required.
Conclusions
As the Mediterranean depletion of biodiversity increases due to urban pressure, forest fires and engulfment in invasive alien species (e.g.
Acknowledgements
Christian Schwarz discussed the subject and provided the reference work of Giglio-Tos (1927). Catarina Nabais and José Alberto Quartau made invaluable suggestions which greatly improved the manuscript and Martin Corley perfectioned it in the best possible way. Finally, two referees and the subject editor cemented the paper's scientific coherence and structure with constructive comments and critics. To all of them the author is really indebted.
References
- The Mediterranean species of the genus Ameles Burmeister, 1838 (Insecta, Mantodea: Amelinae) with a biogeographic and phylogenetic evaluation.Boletín de la Sociedad Entomológica Aragonesa (S.E.A)47:1‑20.
- Contribution to the knowledge of the genus Ameles (Burmeister, 1838) with the description of a new species from Jordan (Insecta Mantodea).Atti Accademia Roveretana degli Agiati a8:173‑197.
- Mantids of the Euro-Mediterranean Area.WBA Handbooks,Verona,240pp.
- New finding of the threatened Apteromantis aptera (Fuente, 1893) in Portugal (Mantodea).Boletin de la Sociedad Entomologica Aragonesa (S. E. A)41:447‑448.
- Sinópsis de los Ortópteros de España y Portugal.Anales de la Sociedad Española de Historia Natural de Madrid5:79‑130.
- Recent range expansion of the Praying mantis, Mantis religiosa Linnaeus (Mantodea; Mantidae) in British Columbia.Journal of the Entomological Society of British Columbia104:73‑80.
- Invasive species are a leading cause of animal extinctions.Trends in ecology and evolution20(3):110.
- Biogeografia de Portugal Continental.Quercetea0:5‑56.
- Mantis movements by night and the interactions of sympatric bats and mantises.Canadian Journal of Zoology74(9):1771‑1774. https://doi.org/10.1139/z96-194
- Handbook of Alien Species in Europe.Springer, Dordrecht,399pp. https://doi.org/10.1007/978-1-4020-8280-1
- Biological Invasions in Europe and the Mediterranean Basin.Dordrecht: Kluwer,463pp. https://doi.org/10.1007/978-94-009-1876-4
- Defensive behaviour in Ghanaian praying mantids.Zoological Journal of the Linnean Society51(1):1‑32. https://doi.org/10.1111/j.1096-3642.1972.tb00771.x
- Mantodea: Gottesanbeterinnen der Welt.Natur und Tier – Verlag,Münster,519pp. [ISBN3-931587-60-6]
- The Ecology of Invasions by Animals and Plants.Methuen & Co. Ltd,181pp. https://doi.org/10.1007/978-1-4899-7214-9
- Fatal attraction: sexually cannibalistic invaders attract naive native mantids.Biology Letters9(6):20130746‑20130746. https://doi.org/10.1098/rsbl.2013.0746
- Orthoptera. Mantidae.Das Tierreich50:1‑707.
- First record of Apteromantis aptera (Fuente, 1894) for Portugal and confirmation of the occurrence of Perlamantis allibertii Guerin-Meneville, 1843 (Dictyoptera, Mantodea).Boletin de la Sociedad Entomologica Aragonesa (S. E. A)35:277‑277.
- Biogeography of Mediterranean Invasions.Cambridge University Press,485pp. https://doi.org/10.1017/cbo9780511525544
- Living Statilia maculata Thunberg (Insecta: Mantodea: Mantidae) and other invertebrates, frequently imported into Dunedin on used cars.The Weta33:17‑19.
- How to know the grasshoppers, crickets, cockroaches and their allies.2.Dover,New York,363pp.
- Heller K-, Bohn H (2013) Mantodea. Fauna Europaea version 2.6. URL: www.faunaeur.org
- Grasping at the routes of biological invasions: a framework for integrating pathways into policy.Journal of Applied Ecology45(2):403‑414. https://doi.org/10.1111/j.1365-2664.2007.01442.x
- Hurd LE (1999) Ecology of praying mantids. In: Prete FR, Wells H, Wells PH, Hurd LE (Eds) The Praying Mantids: Research Perspectives.Johns Hopkins University Press,Baltimore, MD,43-60pp.
- Government funding: Cuts threaten future of R&D in Portugal.Nature506(7487):159. https://doi.org/10.1038/506159d
- Invaded habitats. Chapter 4.BIORISK – Biodiversity and Ecosystem Risk Assessment4:45‑50. https://doi.org/10.3897/biorisk.4.66
- Sphodromantis viridis (Forskal, 1775): New for Portugal and new records of the rare and small mantids Apteromantis aptera (Fuente, 1894) and Perlamantis allibertii Guerin-Meneville, 1843 in the country (Mantodea: Mantidae and Amorphoscelidae).Biodiversity Data Journal2:1037‑1037. https://doi.org/10.3897/BDJ.2.e1037
- Range expansion of an introduced mantid Iris oratoria and niche overlap with a native mantid Stagmomantis limbata (Mantodea: Mantidae).Annals of the Entomological Society of America91(4):422‑429.
- Biotic homogenization: a few winners replacing many losers in the next mass extinction.Trends in Ecology & Evolution14(11):450‑453. https://doi.org/10.1016/s0169-5347(99)01679-1
- McLeod JH (1962) Part I: Biological control of pests of crops, fruit trees, ornamentals and weeds in Canadaup to 1959. In: McLeod JH, McGugan BM, Coppel HC (Eds) A review of the biological control attempts against insects and weeds in Canada. Technical Communication of the CommonwealthInstitute of Biological Control volume 2.Silwood Park, Ascot.
- Sintra Vegetation and Landscape.Quercetea7:65‑81.
- The evolutionary impact of invasive species.Proceedings of the National Academy of Sciences98(10):5446‑5451. https://doi.org/10.1073/pnas.091093398
- Biodiversity hotspots for conservation priorities.Nature203:853‑858.
- On the usefulness of ratios for the identification of some Mediterranean species of the genus Ameles Burmeister, 1838 (Insecta, Mantodea).Zootaxa3259:34‑50.
- Ecological and evolutionary consequences of biotic homogenization.Trends in ecology & evolution19(1):18‑24. https://doi.org/10.1016/j.tree.2003.09.010
- The Praying Mantids.The Johns Hopkins University Press,Baltimore and London,362pp.
- Pathways and vectors of alien arthropods in Europe. Chapter 3.BIORISK – Biodiversity and Ecosystem Risk Assessment4(1):27‑43. https://doi.org/10.3897/biorisk.4.60
- Miomantis caffra, a new mantid record (Mantodea: Mantidae) for New Zealand.New Zealand Entomologist8(1):102‑104. https://doi.org/10.1080/00779962.1984.9722479
- Mantodea (Insecta) with a review of aspects of functional morphology and biology.Fauna of New Zealand19:1‑96.
- La vegetación del sur de Portugal (Sado, Alentejo y Algarve).Itinera Geobotanica3:5‑126.
- Roques A, Rabitsch W, Rasplus J, Lopez-Vaamonde C, Nentwig W, Kenis M (2009) Alien Terrestrial Invertebrates of Europe. In: Hulme PE, W N, Pyšek P, Vilà M (Eds) DAISIE, Handbook of Alien Species in Europe. https://doi.org/10.1007/978-1-4020-8280-1_5
- Landscape disturbance and biodiversity in Mediterranean-type ecosystems.Springer,451pp.
- Global biodiversity scenarios for the year 2100.Science287(5459):1770‑1774.
- Notas entomologicas.Memorias e Estudos do Museu Zoologico da Universidade de Coimbra101:1‑12.
- How well do we understand the impacts of alien species on ecosystem services? A pan-European, cross-taxa assessment.Frontiers in Ecology and the Environment8(3):135‑144. https://doi.org/10.1890/080083
- Alien species in a warmer world: risks and opportunities.Trends in Ecology & Evolution24(12):686‑693. https://doi.org/10.1016/j.tree.2009.06.008
- A review of the research on Canary Islands praying mantises (Mantodea).Zootaxa3797:78‑102.
- Something in the way you move: dispersal pathways affect invasion success.Trends in Ecology & Evolution24(3):136‑144. https://doi.org/10.1016/j.tree.2008.10.007