|
Biodiversity Data Journal :
Taxonomic paper
|
Neoniphon pencei, a new species of holocentrid (Teleostei: Beryciformes) from Rarotonga, Cook Islands
|
Corresponding author:
Academic editor: Rupert Collins
Received: 20 Oct 2014 | Accepted: 22 Jan 2015 | Published: 26 Jan 2015
© 2015 Joshua Copus, Richard Pyle, John Earle
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Copus J, Pyle R, Earle J (2015) Neoniphon pencei, a new species of holocentrid (Teleostei: Beryciformes) from Rarotonga, Cook Islands. Biodiversity Data Journal 3: e4180. https://doi.org/10.3897/BDJ.3.e4180
|
|
Abstract
Neoniphon pencei, n. sp., is described from thirteen specimens, 132-197 mm standard length (SL) collected from mesophotic coral ecosystems (MCEs) at Rarotonga, Cook Islands by divers using mixed-gas closed-circuit rebreathers. It differs from all other species of the genus in number of lateral line scales, scales above and below lateral line, elements of life color, and in COI and cytochrome b DNA sequences. Of the five other known species of Neoniphon, it is most similar to the Indo-Pacific N. aurolineatus and the western Atlantic N. marianus both morphologically and genetically.
Keywords
Holocentridae, Neoniphon, new species, Mesophotic Coral Ecosystems, MCE, Rarotonga, Cook Islands
Introduction
Collections of shore fishes at mesophotic depths (~30-200 m) across the Indo-Pacific are yielding a surprising number of undescribed species (
The genus Neoniphon
Materials and methods
Type specimens of the new species Neoniphon pencei from Rarotonga, Cook Islands have been deposited in the Bernice P. Bishop Museum, Honolulu (BPBM); the California Academy of Sciences, San Francisco (CAS); and the U.S. National Museum of Natural History, Washington, D.C. (USNM).
Measurements and counts given here follow the methods outlined in
Tissue samples were obtained from each of the thirteen individuals of N. pencei collected at Rarotonga, Cook Islands by spear at 90-115 m. Tissue samples were also obtained from twenty-two specimens of the five other species of Neoniphon: N. sammara (n=6) collected from Diego Garcia, British Indian Ocean Territory; N. opercularis (n=2) collected from Moorea, French Polynesia; N. aurolineatus (n=7) collected from Oahu, Hawaii; N. marianus (n=1) collected from the Commonwealth of the Bahamas; and N. argenteus (n=6) collected from the Republic of Kiritimati. Total genomic DNA was extracted from each sample using the 'HotSHOT' protocol (
A DNA barcode (cytochrome c oxidase I; COI) was completed for the holotype and one paratype (BPBM 41196) using the primers from
Taxon treatment
Neoniphon pencei , sp. n.
- ZooBank urn:lsid:zoobank.org:act:43F5CABA-6E4B-42BB-8569-8F93D3502DE9
- Barcode of Life NPE001-14
- GenBank KJ201926
- GenBank KJ188437
-
scientificName: Neoniphon pencei; originalNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; originalNameUsageID: ; namePublishedIn:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; namePublishedInID: ; nameAccordingTo:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; nameAccordingToID: ; acceptedNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; acceptedNameUsageID: ; taxonID: ; scientificNameID: ; parentNameUsageID:b047f156-f8da-4ec6-9f64-87345b68a759; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Pence's Squirrelfish; nomenclaturalCode:ICZN; genus:Neoniphon; specificEpithet:pencei; scientificNameAuthorship:Copus, Pyle & Earle; waterBody:Pacific Ocean; islandGroup:Cook Islands; island:Rarotonga; country:Cook Islands; countryCode:CK; locality:E side; Matavera; off Charles J. Boyle's house; verbatimLocality:Cook Islands; Rarotonga; E side; Matavera; off Charles J. Boyle's house; verbatimDepth:115 m; minimumDepthInMeters:115; maximumDepthInMeters:115; decimalLatitude:-21.223798; decimalLongitude:-159.728123; geodeticDatum:WGS 84; coordinateUncertaintyInMeters:300; georeferenceSources:Google Earth; samplingProtocol:Spear; eventDate:2012-07-02; year:2012; month:7; day:2; habitat:small cave near base of vertical drop-off; individualCount:1; lifeStage:adult; preparations:55% Isopropyl; catalogNumber:41197; recordedBy:David F. Pence; disposition:in collection; associatedSequences: ; identifiedBy:Richard L. Pyle; dateIdentified:2012-08-07; modified:2014-10-09T23:30:00Z; language:en; collectionID: ; institutionCode:BPBM; collectionCode:Fish; basisOfRecord:PreservedSpecimen
-
scientificName: Neoniphon pencei; originalNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; originalNameUsageID: ; namePublishedIn:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; namePublishedInID: ; nameAccordingTo:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; nameAccordingToID: ; acceptedNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; acceptedNameUsageID: ; taxonID: ; scientificNameID: ; parentNameUsageID:b047f156-f8da-4ec6-9f64-87345b68a759; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Pence's Squirrelfish; nomenclaturalCode:ICZN; genus:Neoniphon; specificEpithet:pencei; scientificNameAuthorship:Copus, Pyle & Earle; waterBody:Pacific Ocean; islandGroup:Cook Islands; island:Rarotonga; country:Cook Islands; countryCode:CK; locality:E side; Matavera; off Charles J. Boyle's house; verbatimLocality:Cook Islands; Rarotonga; E side; Matavera; off Charles J. Boyle's house; verbatimDepth:115 m; minimumDepthInMeters:115; maximumDepthInMeters:115; decimalLatitude:-21.223798; decimalLongitude:-159.728123; geodeticDatum:WGS 84; coordinateUncertaintyInMeters:300; georeferenceSources:Google Earth; samplingProtocol:Spear; eventDate:2012-07-03; year:2012; month:7; day:3; habitat:small cave near base of vertical drop-off; individualCount:1; lifeStage:adult; preparations:55% Isopropyl; catalogNumber:41196; recordedBy:David F. Pence; disposition:in collection; associatedSequences:GenBank KJ201926; identifiedBy:Richard L. Pyle; dateIdentified:2012-08-07; modified:2014-10-09T23:30:00Z; language:en; collectionID: ; institutionCode:BPBM; collectionCode:Fish; basisOfRecord:PreservedSpecimen
-
scientificName: Neoniphon pencei; originalNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; originalNameUsageID: ; namePublishedIn:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; namePublishedInID: ; nameAccordingTo:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; nameAccordingToID: ; acceptedNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; acceptedNameUsageID: ; taxonID: ; scientificNameID: ; parentNameUsageID:b047f156-f8da-4ec6-9f64-87345b68a759; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Pence's Squirrelfish; nomenclaturalCode:ICZN; genus:Neoniphon; specificEpithet:pencei; scientificNameAuthorship:Copus, Pyle & Earle; waterBody:Pacific Ocean; islandGroup:Cook Islands; island:Rarotonga; country:Cook Islands; countryCode:CK; locality:E side; Matavera; off Charles J. Boyle's house; verbatimLocality:Cook Islands; Rarotonga; E side; Matavera; off Charles J. Boyle's house; verbatimDepth:115 m; minimumDepthInMeters:115; maximumDepthInMeters:115; decimalLatitude:-21.223798; decimalLongitude:-159.728123; geodeticDatum:WGS 84; coordinateUncertaintyInMeters:300; georeferenceSources:Google Earth; samplingProtocol:Spear; eventDate:2012-07-03; year:2012; month:7; day:3; habitat:small cave near base of vertical drop-off; individualCount:1; lifeStage:adult; preparations:55% Isopropyl; catalogNumber:41196; recordedBy:David F. Pence; disposition:in collection; associatedSequences:GenBank KJ201926; identifiedBy:Richard L. Pyle; dateIdentified:2012-08-07; modified:2014-10-09T23:30:00Z; language:en; collectionID: ; institutionCode:BPBM; collectionCode:Fish; basisOfRecord:PreservedSpecimen
-
scientificName: Neoniphon pencei; originalNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; originalNameUsageID: ; namePublishedIn:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; namePublishedInID: ; nameAccordingTo:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; nameAccordingToID: ; acceptedNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; acceptedNameUsageID: ; taxonID: ; scientificNameID: ; parentNameUsageID:b047f156-f8da-4ec6-9f64-87345b68a759; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Pence's Squirrelfish; nomenclaturalCode:ICZN; genus:Neoniphon; specificEpithet:pencei; scientificNameAuthorship:Copus, Pyle & Earle; waterBody:Pacific Ocean; islandGroup:Cook Islands; island:Rarotonga; country:Cook Islands; countryCode:CK; locality:E side; Matavera; off Charles J. Boyle's house; verbatimLocality:Cook Islands; Rarotonga; E side; Matavera; off Charles J. Boyle's house; verbatimDepth:115 m; minimumDepthInMeters:115; maximumDepthInMeters:115; decimalLatitude:-21.223798; decimalLongitude:-159.728123; geodeticDatum:WGS 84; coordinateUncertaintyInMeters:300; georeferenceSources:Google Earth; samplingProtocol:Spear; eventDate:2012-07-03; year:2012; month:7; day:3; habitat:small cave near base of vertical drop-off; individualCount:1; lifeStage:adult; preparations:55% Isopropyl; catalogNumber:41196; recordedBy:David F. Pence; disposition:in collection; associatedSequences:GenBank KJ201926; identifiedBy:Richard L. Pyle; dateIdentified:2012-08-07; modified:2014-10-09T23:30:00Z; language:en; collectionID: ; institutionCode:BPBM; collectionCode:Fish; basisOfRecord:PreservedSpecimen
-
scientificName: Neoniphon pencei; originalNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; originalNameUsageID: ; namePublishedIn:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; namePublishedInID: ; nameAccordingTo:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; nameAccordingToID: ; acceptedNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; acceptedNameUsageID: ; taxonID: ; scientificNameID: ; parentNameUsageID:b047f156-f8da-4ec6-9f64-87345b68a759; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Pence's Squirrelfish; nomenclaturalCode:ICZN; genus:Neoniphon; specificEpithet:pencei; scientificNameAuthorship:Copus, Pyle & Earle; waterBody:Pacific Ocean; islandGroup:Cook Islands; island:Rarotonga; country:Cook Islands; countryCode:CK; locality:E side; Matavera; off Charles J. Boyle's house; verbatimLocality:Cook Islands; Rarotonga; E side; Matavera; off Charles J. Boyle's house; verbatimDepth:115 m; minimumDepthInMeters:115; maximumDepthInMeters:115; decimalLatitude:-21.223798; decimalLongitude:-159.728123; geodeticDatum:WGS 84; coordinateUncertaintyInMeters:300; georeferenceSources:Google Earth; samplingProtocol:Spear; eventDate:2012-07-03; year:2012; month:7; day:3; habitat:small cave near base of vertical drop-off; individualCount:1; lifeStage:adult; preparations:55% Isopropyl; catalogNumber:41196; recordedBy:David F. Pence; disposition:in collection; associatedSequences:GenBank KJ201926; identifiedBy:Richard L. Pyle; dateIdentified:2012-08-07; modified:2014-10-09T23:30:00Z; language:en; collectionID: ; institutionCode:BPBM; collectionCode:Fish; basisOfRecord:PreservedSpecimen
-
scientificName: Neoniphon pencei; originalNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; originalNameUsageID: ; namePublishedIn:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; namePublishedInID: ; nameAccordingTo:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; nameAccordingToID: ; acceptedNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; acceptedNameUsageID: ; taxonID: ; scientificNameID: ; parentNameUsageID:b047f156-f8da-4ec6-9f64-87345b68a759; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Pence's Squirrelfish; nomenclaturalCode:ICZN; genus:Neoniphon; specificEpithet:pencei; scientificNameAuthorship:Copus, Pyle & Earle; waterBody:Pacific Ocean; islandGroup:Cook Islands; island:Rarotonga; country:Cook Islands; countryCode:CK; locality:E side; Matavera; off Charles J. Boyle's house; verbatimLocality:Cook Islands; Rarotonga; E side; Matavera; off Charles J. Boyle's house; verbatimDepth:115 m; minimumDepthInMeters:115; maximumDepthInMeters:115; decimalLatitude:-21.223798; decimalLongitude:-159.728123; geodeticDatum:WGS 84; coordinateUncertaintyInMeters:300; georeferenceSources:Google Earth; samplingProtocol:Spear; eventDate:2012-07-03; year:2012; month:7; day:3; habitat:small cave near base of vertical drop-off; individualCount:1; lifeStage:adult; preparations:55% Isopropyl; catalogNumber:41196; recordedBy:David F. Pence; disposition:in collection; associatedSequences:GenBank KJ201926; identifiedBy:Richard L. Pyle; dateIdentified:2012-08-07; modified:2014-10-09T23:30:00Z; language:en; collectionID: ; institutionCode:BPBM; collectionCode:Fish; basisOfRecord:PreservedSpecimen
-
scientificName: Neoniphon pencei; originalNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; originalNameUsageID: ; namePublishedIn:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; namePublishedInID: ; nameAccordingTo:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; nameAccordingToID: ; acceptedNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; acceptedNameUsageID: ; taxonID: ; scientificNameID: ; parentNameUsageID:b047f156-f8da-4ec6-9f64-87345b68a759; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Pence's Squirrelfish; nomenclaturalCode:ICZN; genus:Neoniphon; specificEpithet:pencei; scientificNameAuthorship:Copus, Pyle & Earle; waterBody:Pacific Ocean; islandGroup:Cook Islands; island:Rarotonga; country:Cook Islands; countryCode:CK; locality:E side; Matavera; off Charles J. Boyle's house; verbatimLocality:Cook Islands; Rarotonga; E side; Matavera; off Charles J. Boyle's house; verbatimDepth:115 m; minimumDepthInMeters:115; maximumDepthInMeters:115; decimalLatitude:-21.223798; decimalLongitude:-159.728123; geodeticDatum:WGS 84; coordinateUncertaintyInMeters:300; georeferenceSources:Google Earth; samplingProtocol:Spear; eventDate:2012-07-03; year:2012; month:7; day:3; habitat:small cave near base of vertical drop-off; individualCount:1; lifeStage:adult; preparations:55% Isopropyl; catalogNumber:41196; recordedBy:David F. Pence; disposition:in collection; associatedSequences:GenBank KJ201926; identifiedBy:Richard L. Pyle; dateIdentified:2012-08-07; modified:2014-10-09T23:30:00Z; language:en; collectionID: ; institutionCode:BPBM; collectionCode:Fish; basisOfRecord:PreservedSpecimen
-
scientificName: Neoniphon pencei; originalNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; originalNameUsageID: ; namePublishedIn:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; namePublishedInID: ; nameAccordingTo:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; nameAccordingToID: ; acceptedNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; acceptedNameUsageID: ; taxonID: ; scientificNameID: ; parentNameUsageID:b047f156-f8da-4ec6-9f64-87345b68a759; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Pence's Squirrelfish; nomenclaturalCode:ICZN; genus:Neoniphon; specificEpithet:pencei; scientificNameAuthorship:Copus, Pyle & Earle; waterBody:Pacific Ocean; islandGroup:Cook Islands; island:Rarotonga; country:Cook Islands; countryCode:CK; locality:E side; Matavera; off Charles J. Boyle's house; verbatimLocality:Cook Islands; Rarotonga; E side; Matavera; off Charles J. Boyle's house; verbatimDepth:115 m; minimumDepthInMeters:115; maximumDepthInMeters:115; decimalLatitude:-21.223798; decimalLongitude:-159.728123; geodeticDatum:WGS 84; coordinateUncertaintyInMeters:300; georeferenceSources:Google Earth; samplingProtocol:Spear; eventDate:2012-07-03; year:2012; month:7; day:3; habitat:small cave near base of vertical drop-off; individualCount:1; lifeStage:adult; preparations:55% Isopropyl; catalogNumber:41196; recordedBy:David F. Pence; disposition:in collection; associatedSequences:GenBank KJ201926; identifiedBy:Richard L. Pyle; dateIdentified:2012-08-07; modified:2014-10-09T23:30:00Z; language:en; collectionID: ; institutionCode:BPBM; collectionCode:Fish; basisOfRecord:PreservedSpecimen
-
scientificName: Neoniphon pencei; originalNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; originalNameUsageID: ; namePublishedIn:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; namePublishedInID: ; nameAccordingTo:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; nameAccordingToID: ; acceptedNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; acceptedNameUsageID: ; taxonID: ; scientificNameID: ; parentNameUsageID:b047f156-f8da-4ec6-9f64-87345b68a759; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Pence's Squirrelfish; nomenclaturalCode:ICZN; genus:Neoniphon; specificEpithet:pencei; scientificNameAuthorship:Copus, Pyle & Earle; waterBody:Pacific Ocean; islandGroup:Cook Islands; island:Rarotonga; country:Cook Islands; countryCode:CK; locality:E side; Matavera; off Charles J. Boyle's house; verbatimLocality:Cook Islands; Rarotonga; E side; Matavera; off Charles J. Boyle's house; verbatimDepth:115 m; minimumDepthInMeters:115; maximumDepthInMeters:115; decimalLatitude:-21.223798; decimalLongitude:-159.728123; geodeticDatum:WGS 84; coordinateUncertaintyInMeters:300; georeferenceSources:Google Earth; samplingProtocol:Spear; eventDate:2012-07-03; year:2012; month:7; day:3; habitat:small cave near base of vertical drop-off; individualCount:1; lifeStage:adult; preparations:55% Isopropyl; catalogNumber:41196; recordedBy:David F. Pence; disposition:in collection; associatedSequences:GenBank KJ201926; identifiedBy:Richard L. Pyle; dateIdentified:2012-08-07; modified:2014-10-09T23:30:00Z; language:en; collectionID: ; institutionCode:BPBM; collectionCode:Fish; basisOfRecord:PreservedSpecimen
-
scientificName: Neoniphon pencei; originalNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; originalNameUsageID: ; namePublishedIn:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; namePublishedInID: ; nameAccordingTo:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; nameAccordingToID: ; acceptedNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; acceptedNameUsageID: ; taxonID: ; scientificNameID: ; parentNameUsageID:b047f156-f8da-4ec6-9f64-87345b68a759; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Pence's Squirrelfish; nomenclaturalCode:ICZN; genus:Neoniphon; specificEpithet:pencei; scientificNameAuthorship:Copus, Pyle & Earle; waterBody:Pacific Ocean; islandGroup:Cook Islands; island:Rarotonga; country:Cook Islands; countryCode:CK; locality:E side; Matavera; off Charles J. Boyle's house; verbatimLocality:Cook Islands; Rarotonga; E side; Matavera; off Charles J. Boyle's house; verbatimDepth:115 m; minimumDepthInMeters:115; maximumDepthInMeters:115; decimalLatitude:-21.223798; decimalLongitude:-159.728123; geodeticDatum:WGS 84; coordinateUncertaintyInMeters:300; georeferenceSources:Google Earth; samplingProtocol:Spear; eventDate:2012-07-03; year:2012; month:7; day:3; habitat:small cave near base of vertical drop-off; individualCount:1; lifeStage:adult; preparations:55% Isopropyl; catalogNumber:41196; recordedBy:David F. Pence; disposition:in collection; associatedSequences:GenBank KJ201926; identifiedBy:Richard L. Pyle; dateIdentified:2012-08-07; modified:2014-10-09T23:30:00Z; language:en; collectionID: ; institutionCode:BPBM; collectionCode:Fish; basisOfRecord:PreservedSpecimen
-
scientificName: Neoniphon pencei; originalNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; originalNameUsageID: ; namePublishedIn:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; namePublishedInID: ; nameAccordingTo:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; nameAccordingToID: ; acceptedNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; acceptedNameUsageID: ; taxonID: ; scientificNameID: ; parentNameUsageID:b047f156-f8da-4ec6-9f64-87345b68a759; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Pence's Squirrelfish; nomenclaturalCode:ICZN; genus:Neoniphon; specificEpithet:pencei; scientificNameAuthorship:Copus, Pyle & Earle; waterBody:Pacific Ocean; islandGroup:Cook Islands; island:Rarotonga; country:Cook Islands; countryCode:CK; locality:E side; Matavera; off Charles J. Boyle's house; verbatimLocality:Cook Islands; Rarotonga; E side; Matavera; off Charles J. Boyle's house; verbatimDepth:115 m; minimumDepthInMeters:115; maximumDepthInMeters:115; decimalLatitude:-21.223798; decimalLongitude:-159.728123; geodeticDatum:WGS 84; coordinateUncertaintyInMeters:300; georeferenceSources:Google Earth; samplingProtocol:Spear; eventDate:2012-07-03; year:2012; month:7; day:3; habitat:small cave near base of vertical drop-off; individualCount:1; lifeStage:adult; preparations:55% Isopropyl; catalogNumber:41196; recordedBy:David F. Pence; disposition:in collection; associatedSequences:GenBank KJ201926; identifiedBy:Richard L. Pyle; dateIdentified:2012-08-07; modified:2014-10-09T23:30:00Z; language:en; collectionID: ; institutionCode:BPBM; collectionCode:Fish; basisOfRecord:PreservedSpecimen
-
scientificName: Neoniphon pencei; originalNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; originalNameUsageID: ; namePublishedIn:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; namePublishedInID: ; nameAccordingTo:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; nameAccordingToID: ; acceptedNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; acceptedNameUsageID: ; taxonID: ; scientificNameID: ; parentNameUsageID:b047f156-f8da-4ec6-9f64-87345b68a759; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Pence's Squirrelfish; nomenclaturalCode:ICZN; genus:Neoniphon; specificEpithet:pencei; scientificNameAuthorship:Copus, Pyle & Earle; waterBody:Pacific Ocean; islandGroup:Cook Islands; island:Rarotonga; country:Cook Islands; countryCode:CK; locality:E side; Matavera; off Charles J. Boyle's house; verbatimLocality:Cook Islands; Rarotonga; E side; Matavera; off Charles J. Boyle's house; verbatimDepth:115 m; minimumDepthInMeters:115; maximumDepthInMeters:115; decimalLatitude:-21.223798; decimalLongitude:-159.728123; geodeticDatum:WGS 84; coordinateUncertaintyInMeters:300; georeferenceSources:Google Earth; samplingProtocol:Spear; eventDate:2012-07-02; year:2012; month:7; day:2; habitat:small cave near base of vertical drop-off; individualCount:1; lifeStage:adult; preparations:55% Isopropyl; catalogNumber:237596; recordedBy:David F. Pence; disposition:in collection; associatedSequences: ; identifiedBy:Richard L. Pyle; dateIdentified:2012-08-07; modified:2014-10-09T23:30:00Z; language:en; collectionID: ; institutionCode:CAS; collectionCode:Fish; basisOfRecord:PreservedSpecimen
-
scientificName: Neoniphon pencei; originalNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; originalNameUsageID: ; namePublishedIn:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; namePublishedInID: ; nameAccordingTo:Copus, Joshua M., Richard L. Pyle & John L. Earle. 2014. Neoniphon pencei, a new species of holocentrid from Rarotonga, Cook Islands. Biodiversity Data Journal.; nameAccordingToID: ; acceptedNameUsage:Neoniphon pencei Copus, Pyle and Earle, 2014; acceptedNameUsageID: ; taxonID: ; scientificNameID: ; parentNameUsageID:b047f156-f8da-4ec6-9f64-87345b68a759; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Pence's Squirrelfish; nomenclaturalCode:ICZN; genus:Neoniphon; specificEpithet:pencei; scientificNameAuthorship:Copus, Pyle & Earle; waterBody:Pacific Ocean; islandGroup:Cook Islands; island:Rarotonga; country:Cook Islands; countryCode:CK; locality:N side; off Avarua Harbor; verbatimLocality:Cook Islands; Rarotonga; N side; off Avarua Harbor; verbatimDepth:90 m; minimumDepthInMeters:90; maximumDepthInMeters:90; decimalLatitude:-21.198947; decimalLongitude:-159.781353; geodeticDatum:WGS 84; coordinateUncertaintyInMeters:300; georeferenceSources:Google Earth; samplingProtocol:Quinaldine; eventDate:2012-06-22; year:2012; month:6; day:22; individualCount:1; lifeStage:adult; preparations:55% Isopropyl; catalogNumber:431482; recordedBy:John L. Earle; disposition:in collection; otherCatalogNumbers:Formerly BPBM 41195; associatedSequences:GenBank KJ201926; identifiedBy:Richard L. Pyle; dateIdentified:2012-08-07; modified:2014-10-09T23:30:00Z; language:en; collectionID: ; institutionCode:USNM; collectionCode:Fish; basisOfRecord:PreservedSpecimen
-
scientificName: Neoniphon sammara; acceptedNameUsage:N. sammara (Forsskål 1775); parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Sammara squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Indian Ocean; islandGroup:Diego Garcia; country:British Indian Ocean Territory; countryCode:IOT; verbatimLocality:Diego Garcia; samplingProtocol:Spear; year:2002-2011; individualID:NSA128; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Matt Craig; disposition:in collection; associatedSequences:Genbank-KJ188433; identifiedBy:Matt Craig; language:en
-
scientificName: Neoniphon sammara; acceptedNameUsage:N. sammara (Forsskål 1775); parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Sammara squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Indian Ocean; islandGroup:Diego Garcia; country:British Indian Ocean Territory; countryCode:IOT; verbatimLocality:Diego Garcia; samplingProtocol:Spear; year:2002-2011; individualID:NSA129; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Matt Craig; disposition:in collection; associatedSequences:Genbank-KJ188434; identifiedBy:Matt Craig; language:en
-
scientificName: Neoniphon sammara; acceptedNameUsage:N. sammara (Forsskål 1775); parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Sammara squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Indian Ocean; islandGroup:Diego Garcia; country:British Indian Ocean Territory; countryCode:IOT; verbatimLocality:Diego Garcia; samplingProtocol:Spear; year:2002-2011; individualID:NSA130; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Matt Craig; disposition:in collection; associatedSequences:Genbank-KJ188435; identifiedBy:Matt Craig; language:en
-
scientificName: Neoniphon sammara; acceptedNameUsage:N. sammara (Forsskål 1775); parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Sammara squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Indian Ocean; islandGroup:Diego Garcia; country:British Indian Ocean Territory; countryCode:IOT; verbatimLocality:Diego Garcia; samplingProtocol:Spear; year:2002-2011; individualID:NSA131; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Matt Craig; disposition:in collection; associatedSequences:Genbank-KJ188436; identifiedBy:Matt Craig; language:en
-
scientificName: Neoniphon sammara; acceptedNameUsage:N. sammara (Forsskål 1775); parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Sammara squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Indian Ocean; islandGroup:Diego Garcia; country:British Indian Ocean Territory; countryCode:IOT; verbatimLocality:Diego Garcia; samplingProtocol:Spear; year:2002-2011; individualID:NSA132; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Matt Craig; disposition:in collection; associatedSequences:Genbank-KJ188434; identifiedBy:Matt Craig; language:en
-
scientificName: Neoniphon sammara; acceptedNameUsage:N. sammara (Forsskål 1775); parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Sammara squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Indian Ocean; islandGroup:Diego Garcia; country:British Indian Ocean Territory; countryCode:IOT; verbatimLocality:Diego Garcia; samplingProtocol:Spear; year:2002-2011; individualID:NSA133; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Matt Craig; disposition:in collection; associatedSequences:Genbank-KJ188433; identifiedBy:Matt Craig; language:en
-
scientificName: Neoniphon aurolineatus; acceptedNameUsage:N. aurolineatus Liénard 1839; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Yellowstriped squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Pacific Ocean; islandGroup:Hawaii; island:Oahu; country:United States; countryCode:USA; verbatimLocality:Hawaii; samplingProtocol:Spear; year:2012; individualID:NAU1; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Andrew Gray; disposition:in collection; associatedSequences:Genbank-KJ201925; identifiedBy:Andrew Gray; language:en
-
scientificName: Neoniphon aurolineatus; acceptedNameUsage:N. aurolineatus Liénard 1839; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Yellowstriped squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Pacific Ocean; islandGroup:Hawaii; island:Oahu; country:United States; countryCode:USA; verbatimLocality:Hawaii; samplingProtocol:Spear; year:2012; individualID:NAU2; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Andrew Gray; disposition:in collection; associatedSequences:Genbank-KJ201923; identifiedBy:Andrew Gray; language:en
-
scientificName: Neoniphon aurolineatus; acceptedNameUsage:N. aurolineatus Liénard 1839; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Yellowstriped squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Pacific Ocean; islandGroup:Hawaii; island:Oahu; country:United States; countryCode:USA; verbatimLocality:Hawaii; samplingProtocol:Spear; year:2012; individualID:NAU3; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Andrew Gray; disposition:in collection; associatedSequences:Genbank-KJ201924; identifiedBy:Andrew Gray; language:en
-
scientificName: Neoniphon aurolineatus; acceptedNameUsage:N. aurolineatus Liénard 1839; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Yellowstriped squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Pacific Ocean; islandGroup:Hawaii; island:Oahu; country:United States; countryCode:USA; verbatimLocality:Hawaii; samplingProtocol:Spear; year:2012; individualID:NAU4; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Andrew Gray; disposition:in collection; associatedSequences:Genbank-KJ201923; identifiedBy:Andrew Gray; language:en
-
scientificName: Neoniphon aurolineatus; acceptedNameUsage:N. aurolineatus Liénard 1839; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Yellowstriped squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Pacific Ocean; islandGroup:Hawaii; island:Oahu; country:United States; countryCode:USA; verbatimLocality:Hawaii; samplingProtocol:Spear; year:2012; individualID:NAU5; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Andrew Gray; disposition:in collection; associatedSequences:Genbank-KJ201922; identifiedBy:Andrew Gray; language:en
-
scientificName: Neoniphon aurolineatus; acceptedNameUsage:N. aurolineatus Liénard 1839; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Yellowstriped squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Pacific Ocean; islandGroup:Hawaii; island:Oahu; country:United States; countryCode:USA; verbatimLocality:Hawaii; samplingProtocol:Spear; year:2012; individualID:NAU6; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Andrew Gray; disposition:in collection; associatedSequences:Genbank-KJ201925; identifiedBy:Andrew Gray; language:en
-
scientificName: Neoniphon aurolineatus; acceptedNameUsage:N. aurolineatus Liénard 1839; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Yellowstriped squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Pacific Ocean; islandGroup:Hawaii; island:Oahu; country:United States; countryCode:USA; verbatimLocality:Hawaii; samplingProtocol:Spear; year:2012; individualID:NAU7; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Andrew Gray; disposition:in collection; associatedSequences:Genbank-KJ201925; identifiedBy:Andrew Gray; language:en
-
scientificName: Neoniphon opercularis; acceptedNameUsage:Neoniphon opercularis Valenciennes 1831; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Blackfin squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Pacific Ocean; islandGroup:Society; island:Moorea; country:French Polynesia; countryCode:PYF; verbatimLocality:Society; samplingProtocol:Spear; year:2002-2011; individualID:NOP1; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Matt Craig; disposition:in collection; associatedSequences:Genbank-KJ188432; identifiedBy:Matt Craig; language:en
-
scientificName: Neoniphon opercularis; acceptedNameUsage:Neoniphon opercularis Valenciennes 1831; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Blackfin squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Pacific Ocean; islandGroup:Society; island:Moorea; country:French Polynesia; countryCode:PYF; verbatimLocality:Society; samplingProtocol:Spear; year:2002-2011; individualID:NOP2; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Matt Craig; disposition:in collection; associatedSequences:Genbank-KJ188432; identifiedBy:Matt Craig; language:en
-
scientificName: Neoniphon argenteus; acceptedNameUsage:Neoniphon argenteus Valenciennes 1831; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Clearfin squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Pacific Ocean; islandGroup:Line; island:Kiritimati; country:Republic of Kiritimati; countryCode:KIR; verbatimLocality:Line; samplingProtocol:Spear; year:2002-2011; individualID:NAR1; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Matt Craig; disposition:in collection; associatedSequences:Genbank-KJ188431; identifiedBy:Matt Craig; language:en
-
scientificName: Neoniphon argenteus; acceptedNameUsage:Neoniphon argenteus Valenciennes 1831; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Clearfin squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Pacific Ocean; islandGroup:Line; island:Kiritimati; country:Republic of Kiritimati; countryCode:KIR; verbatimLocality:Line; samplingProtocol:Spear; year:2002-2011; individualID:NAR2; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Matt Craig; disposition:in collection; associatedSequences:Genbank-KJ188431; identifiedBy:Matt Craig; language:en
-
scientificName: Neoniphon argenteus; acceptedNameUsage:Neoniphon argenteus Valenciennes 1831; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Clearfin squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Pacific Ocean; islandGroup:Line; island:Kiritimati; country:Republic of Kiritimati; countryCode:KIR; verbatimLocality:Line; samplingProtocol:Spear; year:2002-2011; individualID:NAR3; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Matt Craig; disposition:in collection; associatedSequences:Genbank-KJ188431; identifiedBy:Matt Craig; language:en
-
scientificName: Neoniphon argenteus; acceptedNameUsage:Neoniphon argenteus Valenciennes 1831; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Clearfin squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Pacific Ocean; islandGroup:Line; island:Kiritimati; country:Republic of Kiritimati; countryCode:KIR; verbatimLocality:Line; samplingProtocol:Spear; year:2002-2011; individualID:NAR4; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Matt Craig; disposition:in collection; associatedSequences:Genbank-KJ188431; identifiedBy:Matt Craig; language:en
-
scientificName: Neoniphon argenteus; acceptedNameUsage:Neoniphon argenteus Valenciennes 1831; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Clearfin squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Pacific Ocean; islandGroup:Line; island:Kiritimati; country:Republic of Kiritimati; countryCode:KIR; verbatimLocality:Line; samplingProtocol:Spear; year:2002-2011; individualID:NAR5; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Matt Craig; disposition:in collection; associatedSequences:Genbank-KJ188431; identifiedBy:Matt Craig; language:en
-
scientificName: Neoniphon argenteus; acceptedNameUsage:Neoniphon argenteus Valenciennes 1831; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Clearfin squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Pacific Ocean; islandGroup:Line; island:Kiritimati; country:Republic of Kiritimati; countryCode:KIR; verbatimLocality:Line; samplingProtocol:Spear; year:2002-2011; individualID:NAR6; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Matt Craig; disposition:in collection; associatedSequences:Genbank-KJ188431; identifiedBy:Matt Craig; language:en
-
scientificName: Neoniphon marianus; acceptedNameUsage:Neoniphon marianus Cuvier 1829; parentNameUsage:Neoniphon Castelnau, 1875; higherClassification:Animalia; Deuterostomia; Chordata; Craniata; Gnathostomata; Actinopterygii; Beryciformes; Holocentroidei; Holocentridae; Neoniphon; kingdom:Animalia; phylum:Chordata; class:Actinopterygii; order:Beryciformes; family:Holocentridae; taxonRank:species; vernacularName:Longjaw squirelfish; nomenclaturalCode:ICZN; genus:Neoniphon; waterBody:Atlantic Ocean; islandGroup:Bahamas; island:Bahamas; country:Commonwelth of the Bahamas; countryCode:BHS; verbatimLocality:Bahamas; samplingProtocol:Spear; year:2013; individualID:NMA1; individualCount:1; lifeStage:adult; preparations:DMSO; recordedBy:Matt Craig; disposition:in collection; associatedSequences:Genbank-KJ201921; identifiedBy:Casey Benkwitt; language:en
Description
Dorsal rays XI, 13, the last ray branched to base; anal rays IV,9, the last ray branched to base; principal caudal rays 17, the upper and lower unbranched; upper procurrent caudal rays 7, the first spinous, the last slender and segmented; lower procurrent caudal rays 6, the first 5 spinous, the last slender and segmented; pectoral rays 14, the uppermost rudimentary, the second and lowermost unbranched; pelvic rays I,7; lateral line scales 49 (48-52); scales above lateral line to base of dorsal spines 5; scales below lateral line to origin of anal fin 6 (6-7); oblique rows of scales on cheek 5; vertical row of 9 (8-10) scales on opercle; gill rakers 6+13 (6-7+13); vertebrae 25; body depth 3.2 (2.9-3.3) in SL; head length 2.8 (2.6-2.9) in SL; snout length 3.5 (3.4-3.8) in head length; orbit diameter 3.0 (2.6-3.0) in head length; interorbital width 4.8 (4.1-4.8) in body depth; upper-jaw length 2.3 (2.3-2.6) in head length; preopercular spine 2.2 (2.0-2.9) in orbit diameter; caudal peduncle depth 3.9 (3.7-4.3) in body depth; caudal peduncle length 7.3 (6.5-7.7) in SL, predorsal length 2.7 (2.4-2.7) in SL; preanal length 1.3 (1.2-1.3) in SL; prepelvic length 2.4 (2.4-2.6) in SL; first dorsal spine 3.5 (3.3-4.1) in head length; third dorsal spine longest, 2.3 (2.0-2.8) in head length; first anal spine 29.5 (19.0-29.5) in head length; second anal spine 9.0 (7.2-9.0) in head length; third anal spine 1.3 (1.1-1.3) in head length; fourth anal spine 2.0 (1.7-2.0) in head length; longest anal ray 2.0 (1.9-2.2) in head length; caudal-fin length 5.0 (3.3-5.2) in SL; caudal concavity 2.9 (2.0-2.9) in head length; pectoral-fin length 4.1 (3.6-4.2) in SL; pelvic-spine length 2.3 (2.2-2.5) in head length; pelvic-fin length 4.9 (4.3-4.9) in SL.
Color in life (Figs



Color in alcohol: Body pale yellowish-white. Narrow orange-tan stripes bisecting scales of horizontal scale rows, except for lateral line scale row, the stripes above lateral line faint, barely visible on some specimens. Preopercle white with narrow yellow-tan border. Opercle, nape and interorbital space yellow-tan. Spinous dorsal fin membranes translucent with a white tint. Soft dorsal fin, anal fin pectoral fin and pelvic fin membranes transparent. Caudal fin with orange-tan blotch on upper and lower base, extending faintly on to upper and lower rays. Middle third of caudal fin rays and membranes transparent.
Diagnosis
Dorsal rays XI,13; anal rays IV,9; pectoral rays 14; lateral-line scales 48-52 (usually 49); scales above lateral line to base of dorsal spines 5; scales below lateral line to base of anal fin 6-7; oblique rows of scales on cheek 5; gill rakers 6-7+13 (usually 6+13); body slender, the depth 2.9-3.3 in SL; head length 2.6-2.9 in SL; orbit diameter 2.7-3.0 in head length; interorbital width 4.1-4.8 in body depth; upper jaw length 2.3-2.6 in head; lower jaw strongly protruding; preopercular spine 2.0-2.9 of orbit diameter; first dorsal spine 3.3-4.1 in head length; last dorsal spine shortest; third anal spine the longest, its length 1.1-1.3 in head length; body red with white stripes dorsally, front edge of pelvic and anal fins white, white on base of dorsal fin; reaches 24cm.
Etymology
Named for David F. Pence, Dive Safety Officer for the University of Hawai'i, a member of the deep diving team that discovered this species, in recognition of his efforts to collect the type specimens.
Distribution
All type specimens of N. pencei were collected at Rarotonga, Cook Islands. An individual Neoniphon closely matching the life colors of N. pencei (and different from all other known species) was captured on video by Robert K. Whitton at a depth of 90 m at Moorea, in February 2012 (Fig.
Analysis
Genetic results
After alignment and editing, a 377-bp partial sequence of Cyt b was obtained for all thirty-five Neoniphon samples, resulting in twelve unique haplotypes. All three phylogenetic methods used resulted in congruent tree topologies and are presented as a Maximum Likelihood reconstruction (Fig.
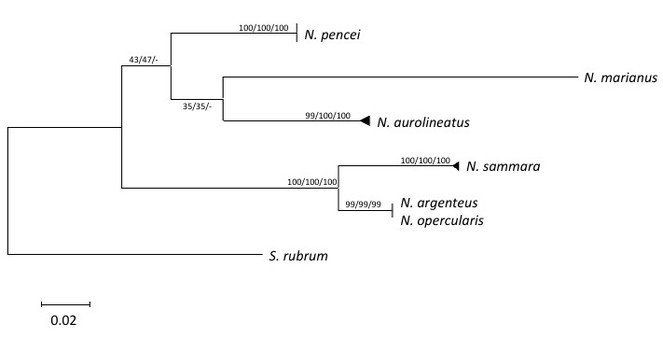
Maximum likelihood phylogenetic reconstruction for the genus Neoniphon based on Cyt b sequences from 35 individuals, yielding 12 unique haplotypes, rooted with Sargocentron rubrum. Branch support values are Maximum Likelihood, Neighbor-Joining, and Maximum Parsimony bootstrap percent values respectively. Triangles at branch termini represent multiple haplotypes; vertical bars at branch termini represent multiple individuals with identical haplotypes.
Discussion
Most recent authors who have reported on Neoniphon (e.g.,
Meristic data of the type specimens of Neoniphon pencei are included in Table
Holo- type |
Paratypes | ||||||||||||
BPBM 41197 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
USNM 431482 |
CAS 237596 |
|
| 175 | 197 | 172 | 170 | 159 | 157 | 162 | 150 | 160 | 160 | 135 | 165 | 132 | |
Dorsal Fin Rays |
XI,13 | XI,13 | XI,13 | XI,13 | XI,13 | XI,13 | XI,13 | XI,13 | XI,13 | XI,13 | XI,13 | XI,13 | XI,13 |
Anal Fin Rays |
IV,9 | IV,9 | IV,9 | IV,9 | IV,9 | IV,9 | IV,9 | IV,9 | IV,9 | IV,9 | IV,9 | IV,9 | IV,9 |
Pectoral Fin Rays |
14 | 14 | 14 | 14 | 14 | 14 | 14 | 14 | 14 | 14 | 14 | 14 | 14 |
Pelvic Fin Rays |
I,7 | I,7 | I,7 | I,7 | I,7 | I,7 | I,7 | I,7 | I,7 | I,7 | I,7 | I,7 | I,7 |
Principal Caudal Rays |
9+8 | 9+8 | 9+8 | 9+8 | 9+8 | 9+8 | 9+8 | 9+8 | 9+8 | dama- ged |
9+8 | 9+8 | 9+8 |
Upper Pro- current Caudal Rays |
7 | 7 | 7 | 7 | 7 | 7 | 7 | 7 | 7 | dama- ged |
7 | 7 | 7 |
Lower Pro- current Caudal Rays |
6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 | dama- ged |
6 | 6 | 6 |
Lateral Line Scales |
49 | 49 | 49 | 49 | 49 | 49 | 49 | 48 | 49 | dama- ged |
49 | 52 | 49 |
Scales Above Lateral Line |
5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 |
Scales Below Lateral Line |
7 | 7 | 6 | 7 | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 7 |
Cheek Scales |
5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 |
Opercle Scales |
9 | 9 | 10 | 10 | 9 | 10 | 8 | 9 | 10 | 10 | 9 | 9 | 9 |
Gill Rakers |
6+13 | 7+13 | 6+13 | 6+13 | 6+13 | 6+13 | 6+13 | 6+13 | 7+13 | 6+13 | 7+13 | 6+13 | 6+13 |
Verte- brate |
25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 |
Proportional measurements of type specimens of Neoniphon pencei as percentages of standard length1, head length2, orbit diameter3, or body depth4.
Holo- type |
Paratypes | ||||||||||||
BPBM 41197 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
BPBM 41196 |
USNM 431482 |
CAS 237596 |
|
Standard length (mm) |
175 | 197 | 172 | 170 | 170 | 157 | 162 | 150 | 160 | 160 | 135 | 165 | 132 |
Body depth1 |
3.15 | 3.30 | 3.04 | 3.06 | 3.09 | 2.90 | 3.16 | 3.06 | 3.02 | 3.06 | 3.16 | 3.08 | 3.03 |
Head length1 |
2.82 | 2.93 | 2.77 | 2.79 | 2.83 | 2.60 | 2.72 | 2.69 | 2.74 | 2.70 | 2.81 | 2.84 | 2.73 |
Snout length2 |
3.54 | 3.60 | 3.70 | 3.49 | 3.69 | 3.54 | 3.72 | 3.78 | 3.66 | 3.43 | 3.76 | 3.63 | 3.66 |
Orbit diameter2 |
3.02 | 2.80 | 2.73 | 2.94 | 2.82 | 3.01 | 2.90 | 2.75 | 2.72 | 2.96 | 2.63 | 2.76 | 2.73 |
Inter- orbital width4 |
4.83 | 4.69 | 4.11 | 4.72 | 4.78 | 4.70 | 4.18 | 4.45 | 4.51 | 4.35 | 4.38 | 4.46 | 4.61 |
Upper- jaw length2 |
2.30 | 2.32 | 2.38 | 2.35 | 2.35 | 2.41 | 2.45 | 2.59 | 2.29 | 2.42 | 2.34 | 2.27 | 2.31 |
Preoper- cular spine3 |
2.16 | 2.00 | 2.39 | 2.44 | 2.66 | 2.86 | 2.34 | 2.53 | 1.95 | broken | 2.61 | 2.00 | 2.54 |
Caudal- peduncle depth4 |
3.90 | 3.92 | 4.04 | 3.83 | 3.79 | 4.00 | 3.73 | 3.77 | 3.93 | dama- ged |
4.28 | 4.12 | 4.17 |
Caudal peduncle length1 |
7.30 | 6.74 | 7.24 | 7.23 | 7.16 | 6.82 | 6.48 | 6.45 | 6.81 | dama- ged |
7.01 | 6.88 | 7.68 |
Predorsal length1 |
2.66 | 2.72 | 2.52 | 2.64 | 2.54 | 2.43 | 2.58 | 2.53 | 2.43 | 2.46 | 2.53 | 2.61 | 2.52 |
Preanal length1 |
1.32 | 1.34 | 1.26 | 1.34 | 1.29 | 1.27 | 1.31 | 1.21 | 1.30 | 1.25 | 1.23 | 1.40 | 1.31 |
Prepelvic length1 |
2.44 | 2.48 | 2.51 | 2.48 | 2.64 | 2.43 | 2.57 | 2.49 | 2.60 | 2.46 | 2.52 | 2.62 | 2.48 |
First dorsal spine2 |
3.50 | 4.14 | 3.99 | broken | 3.66 | 4.09 | 3.65 | 3.54 | 3.86 | 3.65 | 3.45 | 3.84 | 3.33 |
Longest dorsal spine2 |
3.34 | 2.44 | 2.30 | 2.37 | 2.31 | 2.37 | 2.25 | 2.03 | 2.40 | 2.68 | 2.24 | 2.85 | 2.21 |
First anal spine2 |
29.5 | 25.0 | 25.2 | 23.2 | 27.4 | 23.6 | 23.2 | 25.2 | 26.0 | 22.3 | 24.9 | 25.7 | 19.1 |
Second anal spine2 |
9.00 | 8.17 | 8.09 | 7.24 | 8.52 | 8.25 | 8.26 | 7.84 | 8.29 | 7.66 | 7.88 | 7.95 | 7.34 |
Third anal spine2 |
1.34 | 1.24 | 1.21 | 1.20 | 1.25 | 1.22 | 1.26 | 1.16 | 1.13 | 1.17 | 1.13 | 1.18 | 1.16 |
Fourth anal spine2 |
1.98 | 1.97 | 1.80 | 1.89 | 1.89 | 1.93 | 2.03 | 2.00 | 1.74 | 1.99 | 1.71 | 1.89 | 1.88 |
Longest anal ray2 |
2.05 | 2.05 | 1.92 | 1.96 | 2.0 | 2.09 | 2.13 | 2.12 | 1.94 | 2.21 | 1.89 | 2.12 | 1.97 |
Caudal- fin length1 |
5.00 | 5.18 | 4.30 | 4.25 | 4.25 | 3.65 | 3.77 | 3.33 | 3.76 | 4.00 | 4.50 | 4.71 | 3.53 |
Caudal concavity2 |
2.88 | 2.54 | 2.23 | 2.30 | 2.47 | 1.98 | 2.20 | 2.45 | 2.21 | dama- ged |
2.78 | 2.90 | 2.16 |
Pectoral- fin length1 |
4.12 | 4.08 | 4.14 | 4.20 | 4.0 | 3.63 | 3.90 | 3.92 | 4.05 | 3.79 | 3.88 | 4.29 | 4.11 |
Pelvic- spine length2 |
2.32 | 2.26 | 2.21 | 2.26 | 2.31 | 2.51 | 2.33 | 2.23 | 2.29 | 2.37 | 2.29 | 2.47 | 2.43 |
Pelvic- fin length1 |
4.93 | 4.83 | 4.74 | 4.42 | 4.59 | 4.30 | 4.63 | 4.41 | 4.44 | 4.54 | 4.58 | 4.93 | 4.57 |
Comparison of selected morphological characters for species of Neoniphon. Data for N. argenteus, N. aurolineatus (as Flammeo scythrops), N. sammara, and N. opercularis are from
Characters that differ from N. pencei are shown in bold. 1 as a proportion of Standard Length; 2 as a a proportion of orbit diameter; 3 as a proportion of head length.
| Character | N. pencei | N. argenteus | N. aurolineatus | N. marianus | N. opercularis | N. sammara |
|---|---|---|---|---|---|---|
| Head Length1 | 2.6-2.9 | 2.7-3.4 | 2.8-3.1 | 2.6-2.9 | 2.9-3.1 | 2.9-3.2 |
| Snout Length2 | 1.2-1.42 | 9.2-10.61 | 1.2-1.6 | 1.2-1.5 | 9.5-10.61 | 0.8-1.0 | 1.1-1.3 |
| Orbit Diameter3 | 2.6-3.03 | 7.4-8.51 | 2.4-3.0 | 2.5-2.9 | 6.5-8.21 | 3.0-3.5 | 2.5-3.0 |
| Interorbital Width2 | 1.7-1.92 | 12.5-15.41 | 1.2-1.7 | 1.6-2.1 | 11.5-14.81 | 1.2-1.5 | 1.3-1.6 |
| Upper-jaw Length3 | 2.3-2.63 | 6.3-7.01 | 2.2-2.3 | 2.0-2.3 | 5.9-6.71 | 2.1-2.2 | 2.2-2.4 |
| First Dorsal-spine Length3 | 3.3-4.13 | 9.1-12.11 | 2.4-3.1 | 3.2-4.5 | 141 | 3.1.-3.6 | 2.2-3.0 |
| Third Anal-spine Length3 | 1.1-1.33 | 3.1-3.81 | 1.4-1.6 | 1.4-1.6 | 3.4-4.41 | 1.5-1.9 | 1.1-1.5 |
| Fourth Anal-spine Length3 | 1.7-2.0 | 1.9-2.4 | 2.1-2.7 | - | 2.0-2.7 | 1.8-2.2 |
| Dorsal-fin soft rays | 13 | 11-13 | 12-13 | 12-13 | 13 | 11-12 |
| Pectoral-fin Rays | 14 | 12-13 | 14 | 14 | 13-14 | 13-14 |
| Lateral-line Scales | 48-52 | 38-43 | 44-46 | 46-47 | 38-40 | 39-43 |
| Scales Above Lateral Line | 5 | 2.5 | 3.5 | 3.5 | 2.5 | 2.5 |
| Scales Below Lateral Line | 6-7 | 7-8 | 8-9 | 8 | 8 | 8 |
| Gill Rakers | 19-20 | 12-19 | 15-17 | 18-19 | 17-19 | 13-20 |
Neoniphon pencei appears most similar to N. aurolineatus and N. marianus, based on having the the fewest number of differences in morphometrics, greatest genetic similarity, and most similar aspects of life coloration with these two species. It is also similar to N. aurolineatus in the depth and habitat it occupies. However, the differences between N. pencei and these two species as noted above clearly warrant recognition of N. pencei as a distinct species. A more comprehensive phylogenetic analysis of the species of Neoniphon and related genera based on both morphology and genetics (with verified voucher specimens) is beyond the scope of this work.
Acknowledgements
We are especially grateful to Charles "Chip" Boyle for logistical support in the field, and to David F. Pence for collecting most of the type specimens of the new species. Radiographs were produced by Loreen R. O'Hara, and curatorial support was provided by Arnold Y. Suzumoto. We would also like to thank Casey Benkwitt, Tye Kindinger, Alix Davis, Lillina Tuttle, Mark Hixon, Matthew Craig, Athlene Clarke, Richard Coleman, Brian Bowen, John E. Randall, Brian D. Greene and Robert K. Whitton for their help on various various aspects of this work. We would also like to thank the reviewers for their invaluable comments.
References
- Genetic identification and color descriptions of early life-history stages of Belizean Phaeoptyx and Astrapogon (Teleostei: Apogonidae) with comments in identification of adult Phaeoptyx.Zootaxa2008:1‑22.
- Evolution and biogeography of marine angelfishes (Pisces: Pomacanthidae).Molecular Phylogenetics and Evolution33:140‑155. https://doi.org/10.1016/j.ympev.2004.04.015
- Researches on the fishes of Australia. Philadelphia Centennial Expedition of 1876. Intercolonial Exhibition Essays, 1875-6.Intercolonial Exhibitions,Melbourne,1-52pp.
- Suite du Livre troisième. Des percoïdes à dorsale unique à sept rayons branchiaux et à dents en velours ou en cardes.Tome troisième.F.G. Levrault,Paris,i-xxviii, [1-2], 1-500, pls. 41-71pp. [InFrench].
- Livre septième. Des squamipennes. Livre huitième. Des poissons à pharyngiens labyrinthiformes.Tome septième.F. G. Levrault,Paris,i-xxix, 1-531, pls. 170-208pp. [InFrench].
- jModelTest 2: more models, new heuristics and parallel computing.Nature Methods9(8):772‑772. https://doi.org/10.1038/nmeth.2109
- Molecular phylogenetics of squirrelfishes and soldierfishes (Teleostei: Beryciformes: Holocentridae): Reconciling more than 100 years of taxonomic confusion.Molecular Phylogenetics and Evolution65(2):727‑738. [InEnglish]. https://doi.org/10.1016/j.ympev.2012.07.020
- Geneious Pro.5.6.6.Biomatters. Release date:2012-10-15. URL: http://www.geneious.com/
- Confidence limits on phylogenies: an approach using the bootstrap.Evolution39:783‑791. https://doi.org/10.2307/2408678
- Molecular phylogenetics of the butterflyfishes (Chaetodontidae): Taxonomy and biogeography of a global coral reef fish family.Molecular Phylogenetics and Evolution45:50‑68. https://doi.org/10.1016/j.ympev.2007.05.018
- Descriptiones animalium avium, amphibiorum, piscium, insectorum, vermium; quae in itinere orientali observavit Petrus Forskål. Post mortem auctoris edidit Carsten Niebuhr. Adjuncta est materia medica kahirina atque tabula maris Rubri geographica.Mölleri apud Heineck et Faber,Hauniæ [= Copenhagen],1-20, i-xxxiv, 1-164pp.
- New, little known and typical berycoid fishes.Proceedings of the Academy of Natural Sciences of Philadelphia56:222‑238. [InEnglish].
- Greenfield DW (2003) Holocentridae. In: Carpenter KE (Ed.) The living marine resources of the Western Central Atlantic. Volume 2: Bony fishes part 1 (Acipenseridae to Grammatidae).2.FAO,Rome,602-1373pp. [InEnglish].
- A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood.Systematic Biology52:696‑704. [InEnglish]. https://doi.org/10.1080/10635150390235520
- Catalogue of the acanthopterygian fishes in the collection of the British Museum. Gasterosteidae, Berycidae, Percidae, Aphredoderidae, Pristipomatidae, Mullidae, Sparidae.1.Taylor & Francis,London,i-xxxi + 1-524pp. [InEnglish].
- Descriptions of new genera and species of fishes from the Hawaiian Islands.Bulletin of the United States Fish Commission22:161‑208. [InEnglish].
- The fishes of North and Middle America: a descriptive catalogue of the species of fish-like vertebrates found in the waters of North America north of the Isthmus of Panama. Part III.Bulletin of the United States National Museum47:2183a‑3136. [InEnglish].
- Beryciform fishes of the world ocean.VNIRO Publishing,368pp.
- Species composition and distribution of holocentrids in the oceans of the world (Holocentridae, Beryciformes).Journal of Ichthyology38(2):170‑189. [InEnglish].
- Liénard E (1839) Poissons. In: Bouton L (Ed.) Dixième rapport annuel sur les travaux de la Société d’Histoire Naturelle de l’île Maurice.10.Imprimeire du Cerneen,l’île Maurice.
- Method for isolation of PCR-ready genomic DNA from zebrafish tissues.BioTechniques43:610‑614. https://doi.org/10.2144/000112619
- Assessing undiscovered fish biodiversity on deep coral reefs using advanced self-contained diving technology.Marine Technology Society Journal34(4):82‑91. [InEnglish]. https://doi.org/10.4031/MTSJ.34.4.11
- Revision of the Indo-Pacific squirrelfishes (Beryciformes: Holocentridae: Holocentrinae) of the genus Sargocentron, with descriptions of four new species.Indo-Pacific Fishes28:1‑105. [InEnglish].
- A review of the squirrelfishes of the subfamily Holocentrinae from the western Indian Ocean and Red Sea.Ichthyological Bulletin of the J. L. B. Smith Institute of Ichthyology49:1‑27.
- Randall JE, Greenfield DW (1999) Family Holocentridae. In: Carpenter KE, Niem VE (Eds) Species identification guide for fisheries purposes. The living marine resources of the western central Pacific. Bony fishes part 2 (Mugilidae to Carangidae).2.FAO,Rome,2069-2790pp. [InEnglish].
- Randall JE, Heemstra PC (1986) Holocentrdae. In: Smith MM, Heemstra PC (Eds) Smiths' Sea Fishes.Macmillan South Africa,Johannesburg,1047pp. [InEnglish].
- Chaetodontoplus poliourus, A new angelfish (Perciformes: Pomacanthidae) from the tropical western Atlantic.The Raffles Bulletin of Zoology57(2):511‑520.
- Mitochondrial DNA and color pattern variation in three western Atlantic Halichoeres (Labridae), with the revalidation of two species.Copeia4:770‑782. https://doi.org/10.1643/CG-04-106
- Historical biogeography and speciation in the reef fish genus Haemulon (Teleostei: Haemulidae).Molecular Phylogenetics and Evolution48:918‑928. https://doi.org/10.1016/j.ympev.2008.05.024
- Satapoomin U (2009) Holocentridae. In: Kimura S, Satapoomin U, Matsuura K (Eds) Fishes of Andaman Sea, west coast of southern Thailand.National Museum of Nature and Science,Tokyo,346pp.
- Review of the squirrelfishes (subfamily Holocentrinae: order Beryciformes) of Japan, with a description of a new species.Japanese Journal of Ichthyology26(2):109‑147. [InEnglish].
- Phylogenetic relations among percid fishes as inferred from mitochondrial cytochrome b DNA sequence data.Molecular Phylogenetics and Evolution10:343‑353. https://doi.org/10.1006/mpev.1998.0542
- Unusually large mitochondrial variation in populations of the blue tit, Parus caeruleus.Molecular Ecology1:27‑36. [InEnglish]. https://doi.org/10.1111/j.1365-294X.1992.tb00152.x
- MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony method.5.2.2.Molecular Biology and Evolution. URL: www.megasoftware.net
- Western Atlantic species of the genus Holocentrus.Fieldiana Zoology37(4):91‑119. [InEnglish]. https://doi.org/10.5962/bhl.title.2972
- Woods L, Sonoda PM (1973) Order Berycomorphi (Beryciformes). Fishes of the western North Atlantic: Halosuriforms, killfishes, squirrelfishes and other beryciforms, stephanoberyciforms, grenadiers.Part 6.The Sears Foundation for Marine Research, Yale University,New Haven, Connecticut,1-698pp. [InEnglish].