|
Biodiversity Data Journal :
Taxonomic Paper
|
|
Corresponding author: Hendrik Freitag (hfreitag@ateneo.edu)
Academic editor: David Bilton
Received: 21 Oct 2020 | Accepted: 28 Dec 2020 | Published: 12 Jan 2021
© 2021 Hendrik Freitag, Rick de Vries, Marta Paterno, Simone Maestri, Massimo Delledonne, Cameron Thompson, Helena Lamed, Rebekah Lambert, Michael Fox, Mariela Gonzalez, Emmanuel Delocado, Marc Sabordo, Clister Pangantihon, Iva Njunjić
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Freitag H, de Vries R, Paterno M, Maestri S, Delledonne M, Thompson CG, Lamed H, Lambert R, Fox MF, Gonzalez MC, Delocado ED, Sabordo MR, Pangantihon CV, Njunjić I (2021) Hydraena (s.str.) dinarica, new species (Coleoptera: Hydraenidae) along with further records of Hydraena spp. from Durmitor National Park, Montenegro and comments on the DNA barcoding problem with the genus. Biodiversity Data Journal 9: e59892. https://doi.org/10.3897/BDJ.9.e59892
|
|
Abstract
Background
Long-palped Water Beetles were collected during a taxon expedition in Montenegro which involved citizen scientists, students and taxonomists. The material was collected from springs, brooks, fens and the Tara River, at altitudes between 600 m and 1450 m above sea level, using fine-meshed hand-nets and by manual checking of submerged substrates. The morphological species delimitation was supplemented and congruent with mtDNA sequences mainly obtained in the field using the newly-developed MinION-based ONTrack pipeline.
New information
The new species Hydraena dinarica Freitag & de Vries, sp. n. from Durmitor Mt. is described, illustrated and compared in detail to closely-related congeners of the H. saga d'Orchymont, 1930/H. emarginata Rey, 1885 species complex. Five additional species and female specimens of two unidentified morphospecies of the genus were also recorded in the vicinity of Durmitor National Park. New records and the first DNA barcodes for Hydraena biltoni Jäch & Díaz, 2012 (endemic to Montenegro) and H. morio Kiesenwetter, 1849 are provided. Further records of H. nigrita Germar, 1824, H. minutissima Stephens, 1829, H. subintegra Ganglbauer, 1901 and females of two unidentified morphospecies are commented upon. The resulting inter- and intraspecific genetic distances and some observations of low or zero sequence divergence between recently-diverged species of Hydraena Kugelann, 1794 are briefly discussed.
Keywords
citizen science, MinION sequencing, minute moss beetle, Palaearctic region, Taxon Expeditions, Durmitor Mt.
Introduction
The Long-palped Water Beetles of the genus Hydraena, originally described by
The subsequent identification of the collected material at the improvised field laboratory (Fig.


Materials and methods
Specimens were collected in a microhabitat-specific approach (
Pre-sorting and genus-level identification were performed by taxon expedition participants mentored by the first author (Fig.
Detailed examination and digital imaging of dissected parts was done using an Olympus CX21 microscope equipped with a DinoEye Eyepiece camera. Habitus photographs were taken under a Zeiss Axio Zoom V 16 microscope with a Canon 5D Mark II SLR attached to the microscope. Images were captured at various focus planes and subsequently stacked using the Helicon Focus software. Genital drawings were compiled after their photographs by vector graphic tools in CorelDRAW v.10.0 software, but in direct comparison with the actual genitals mounted on slides.
After removal of diagnostic parts (glued on entomological cards), the entire remaining specimen of each initially-recognised morphospecies and some unidentifiable female specimens underwent DNA isolation, amplification, sequencing and processing of the 5′-end of the mitochondrial cytochrome oxidase I (COI) gene as described in
The DNA sequence alignment, which also included available standard barcodes of related Hydraena species of the region of
A statistical parsimony haplotype network was constructed by TCS1.21 (
DNA sequences were submitted to International Nucleotide Sequence Database Collaboration (INSDC) through GenBank, as well as to the Barcode of Life Data System (BOLD) under project TXEX.
The type labels of the new species are literally quoted from the specimen’s label under 'bibliographicCitation'. Back slashes indicate the next line in the label.
Taxon treatments
Hydraena (Hydraena) biltoni
-
locationID: MNE21c; continent:Europe; waterBody:small Black Lake tributary; country:MONTENEGRO; municipality:Žabljak; locality:Durmitor National Park; verbatimLocality:small Black Lake tributary creek near war monument, pine forest, pebble in shallow run; verbatimElevation:1435 m; verbatimCoordinates:; eventDate:
43 08 57N; 19 05 42E 2019-07-16; individualID:H69; sex:3 males, 2 females; lifeStage:adults; recordedBy:Hendrik Freitag, Michael F. Fox, Rebekah Lambert; associatedSequences:GenBank: MT784158.1; identifiedBy:Hendrik Freitag, Rick De Vries, Cameron G. Thompson, Helena Lamed, Rebekah Lambert, Michael F. Fox, Mariela Gonzalez, Clister V. Pangantihon; institutionCode:CFM, ZMB; collectionCode:Coleoptera; basisOfRecord:Dried specimens; informationWithheld:MONTENEGRO: Žabljak, Durmitor N.P., small Black Lake tributary creek near war monument, pine forest, pebble in shallow run, 1435 m a.s.l., 43 08 57N 19 05 42E, 16 July 2019, leg M.F. Fox, R. Lambert, H. Freitag (MNE21c)
Distribution
Hydraena biltoni (Fig.

Remarks
We provide the first standard barcode for the species. It varies by only 0.3% from that of the closest congener H. morio Kiesenwetter, 1849 (Suppl. material
Habitat
The specimens were collected in a very small creek, flowing through pine forest and a wet meadow. Bottom pebbles, mixed with CPOM, in moderately fast flowing, shallow portions of the creek, were their microhabitat.
Hydraena (Hydraena) minutissima
-
locationID: MNE17b; continent:Europe; waterBody:Shallow littoral, pool with pebbles; country:Montenegro; municipality:Žabljak; locality:Tara River; verbatimLocality:Tara River, near Bijela Stijena, shallow littoral pool with pebbles; verbatimElevation:600 m; verbatimCoordinates:; eventDate:
43 13 23N; 19 09 57E 2019-07-12; individualID:H60; sex:2 males, 5 females; lifeStage:adults; recordedBy:Hendrik Freitag, Clister V. Pangantihon; associatedSequences:GenBank: MT784159.; identifiedBy:Hendrik Freitag, Rick De Vries, Cameron G. Thompson, Helena Lamed, Rebekah Lambert, Michael F. Fox, Mariela Gonzalez, Clister V. Pangantihon; institutionCode:CFM, ZMB; collectionCode:Coleoptera; basisOfRecord:Dried specimens
Distribution
Hydraena minutissima (Fig.
Habitat
The species was collected in the shallow littoral of the Tara River with pebble deposits on bedrock. The microhabitat was not exposed to strong currents during the time of collection and filamentous algae were partly growing on the surrounding exposed rocks.
Hydraena morio
-
locationID: MNE17b; continent:Europe; waterBody:Small littoral pool with pebbles; country:Montenegro; municipality:Žabljak; locality:Tara River; verbatimLocality:Tara River, near Bijela Stijena, shallow littoral pool with pebbles; verbatimElevation:600 m; verbatimCoordinates:; eventDate:
43 13 23N; 19 09 57E 2019-07-12; individualID:H62, H64, H70, H71; sex:7 males, 2 females; lifeStage:adults; recordedBy:Hendrik Freitag, Clister V. Pangantihon; associatedSequences:GenBank: MT784154.1, MT784155.1, MT784156.1, MT784157.1; identifiedBy:Hendrik Freitag, Rick De Vries, Cameron G. Thompson, Helena Lamed, Rebekah Lambert, Michael F. Fox, Mariela Gonzalez, Clister V. Pangantihon; collectionCode:Coleoptera; basisOfRecord:Dried specimen
Distribution
Hydraena morio (Fig.
Remarks
We provide here the first COI 5′-end sequences (Folmer Region) of the species. See also remarks on H. biltoni.
Habitat
For notes on the habitat, see Hydraena minutissima.
Hydraena (Hydraena) nigrita
-
locationID: MNE17b; continent:Europe; waterBody:Shallow littoral pool with pebbles; country:Montenegro; municipality:Žabljak; locality:Tara River; verbatimLocality:Tara River, near Bijela Stijena, shallow littoral pool with pebbles; verbatimElevation:600 m; verbatimCoordinates:; eventDate:
43 13 23N; 19 09 57E 2019-07-12; individualID:H61; sex:2 males; lifeStage:adults; recordedBy:Hendrik Freitag, Clister V. Pangantihon; associatedSequences:GenBank: MT784150.1; identifiedBy:Hendrik Freitag, Rick De Vries, Cameron G. Thompson, Helena Lamed, Rebekah Lambert, Michael F. Fox, Mariela Gonzalez, Clister V. Pangantiho; institutionCode:CFM, ZMB; collectionCode:Coleoptera; basisOfRecord:Dried specimen
Distribution
Hydraena nigrita (Fig.
Habitat
For notes on the habitat, see Hydraena minutissima.
Hydraena (Hydraena) spp.
-
locationID: MNE10l; continent:Europe; waterBody:Fen meadow near Durmitor N.P entrance; country:Montenegro; municipality:Žabljak; locality:Durmitor N.P.; verbatimLocality:Fen meadow near Durmitor N.P entrance, littoral pool; verbatimElevation:1425 m; verbatimCoordinates:; eventDate:
43 06 08N; 19 10 56E 2019-07-10; individualID:H65; sex:3 females; lifeStage:adults; recordedBy:Hendrik Freitag; associatedSequences:GenBank: MT784151.1; identifiedBy:Hendrik Freitag, Rick De Vries, Cameron G. Thompson, Helena Lamed, Rebekah Lambert, Michael F. Fox, Mariela Gonzalez, Clister V. Pangantihon; identificationRemarks:“cf. britteni”; institutionCode:CFM; collectionCode:Coleoptera; basisOfRecord:Dried specimen -
locationID: MNE13k; continent:Europe; waterBody:fen meadow near Dobri Nugo; country:Montenegro; municipality:Žabljak; locality:Dobri Nugo; verbatimLocality:Fen meadow near Dobri Nugo, partly subterranean water flow, slow run; verbatimElevation:1392 m; verbatimCoordinates:; eventDate:
43 09 15N; 19 06 06E 2019-07-11; individualID:H66; sex:2 females; lifeStage:adults; recordedBy:Hendrik Freitag; associatedSequences:GenBank: MT784152.1; identifiedBy:Hendrik Freitag, Rick De Vries, Cameron G. Thompson, Helena Lamed, Rebekah Lambert, Michael F. Fox, Mariela Gonzalez, Clister V. Pangantihon; identificationRemarks:"sp. female"; institutionCode:CFM; collectionCode:Coleoptera; basisOfRecord:Dried specimen
Remarks
The samples cluster with Hydraena britteni Joy, 1907, originally described from England and Ireland (
Habitat
The specimens were collected from fen-like meadows, one (MNE10) densely vegetated with sedge and horsetail, the other (MNE13) additionally with limestone boulders and gravel densely covered with mosses. In both sites, a creek with clear brownish water, rich in humins, was passing the fens and provides continuous water inputs.
Hydraena subintegra
-
locationID: MNE17c; continent:Europe; waterBody:Black Lake tributary creek; country:Montenegro; municipality:Žabljak; locality:Durmitor N.P.; verbatimLocality:Tara River near Bijela Stijena, littoral, run with pebble; verbatimElevation:600 m; verbatimCoordinates:; eventDate:
43 13 23N; 19 09 57E 2019-07-12; individualID:H63; sex:1 male, 1 female; lifeStage:adults; recordedBy:Hendrik Freitag & Clister Pangantihon; associatedSequences:GenBank: MT784149.; identifiedBy:Hendrik Freitag, Rick De Vries, Cameron G. Thompson, Helena Lamed, Rebekah Lambert, Michael F. Fox, Mariela Gonzalez, Clister V. Pangantihon; institutionCode:CFM; collectionCode:Coleoptera; basisOfRecord:Dried specimens -
locationID: MNE20c/f/h; continent:Europe; waterBody:Black Lake tributary creek; country:Montenegro; municipality:Žabljak; locality:Durmitor N.P.; verbatimLocality:Black Lake tributary creek near old watermill, pine forest; verbatimElevation:1450 m; verbatimCoordinates:; eventDate:
43 09 09N; 19 05 22E 2019-07-16; individualID:H63; sex:1 male, 1 female; lifeStage:adults; recordedBy:Helena Lamed, Mariela Gonzales, Rebekah Lambert, Michael F. Fox, Clister V. Pangantihon; identifiedBy:Hendrik Freitag, Rick De Vries, Cameron G. Thompson, Helena Lamed, Rebekah Lambert, Michael F. Fox, Mariela Gonzalez, Clister V. Pangantihon; institutionCode:NMW, ZMB; collectionCode:Coleoptera; basisOfRecord:Dried specimens
Distribution
The species is distributed in an area between the Adriatic and Black Seas, including the Dinaric Alps (
Remarks
The taxonomy of this species of the "Haenydra" lineage is not yet finally resolved. Three slightly varying morphs are recognised. Our specimens (Fig.
Habitat
All specimens were collected from moderately fast flowing, shallow water, but on varying substrates, including submerged wood, grass bunches and pebble.
Hydraena (Hydraena) dinarica , sp. n.
- ZooBank urn:lsid:zoobank.org:act:E5A0DED7-16AC-4B94-B9B1-72000874197D
- Barcode of Life TXEX049-20
-
locationID: MNE18; continent:Europe; waterBody:Skakala stream; country:Montenegro; municipality:Žabljak; locality:Durmitor, Peradova gora; verbatimElevation:1220 m; locationRemarks:cold water karst creek with predominant flow subsurface; verbatimCoordinates:; samplingProtocol:
43 09 54N; 18 59 59E Manual collection by hand-net from bottom substrates; eventDate:2019-07-13; sex:male; lifeStage:adult; recordedBy:Hendrik Freitag, Clister V. Pangantihon; identifiedBy:Hendrik Freitag; type:Dried specimen; bibliographicCitation:MONTENEGRO: Durmitor, Peradova gora, \ Skakala stream, ca. 1220 m asl., \ 43°09′54″N 18°59′59″E, 13 July 2019 \ leg. H. Freitag & C.V. Pangantihon (MNE18); institutionCode:NMW; basisOfRecord:Dried specimen; informationWithheld:Terminal parts of abdomen, aedeagus and right foretasus (broken off) glued separately on to same entomological card along with holotype specimen.
-
locationID: MNE18; continent:Europe; waterBody:Skakala stream; country:Montenegro; municipality:Žabljak; locality:Durmitor, Peradova gora; verbatimElevation:1220 m; locationRemarks:cold water karst creek with predominant flow subsurface; verbatimCoordinates:; samplingProtocol:
43 09 54N; 18 59 59E Manual collection by hand-net from bottom substrates; eventDate:2019-07-13; individualID:H67, H68; sex:14 males, 11 females; lifeStage:adults; recordedBy:Hendrik Freitag, Clister V. Pangantihon; associatedSequences:GenBank: MT784148.1; identifiedBy:Hendrik Freitag, Rick De Vries, Cameron G. Thompson, Helena Lamed, Rebekah Lambert, Michael F. Fox, Mariela Gonzalez, Clister V. Pangantihon; type:Dried specimens; bibliographicCitation:MONTENEGRO: Durmitor, Peradova gora, \ Skakala stream, ca. 1220 m asl., \ 43°09′54″N 18°59′59″E, 13 July 2019 \ leg. H. Freitag & C.V. Pangantihon (MNE18); institutionCode:CFM, NMW, SMTD, ZMB; collectionCode:Coleoptera -
continent: Europe; country:Montenegro; municipality:Žabljak; locality:Durmitor, Zeleni Vir; samplingProtocol:Manual collection; eventDate:2002-08-02; sex:1 female; lifeStage:adult; recordedBy:Vladimir Pešić; identifiedBy:Manfred A. Jäch; type:Dried specimen; institutionCode:NMW; collectionCode:Coleoptera
Description
Habitus as in Fig.

Pronotum broadly subhexagonal, moderately wider than long; anterior and posterior margins slightly concave; anterior and posterior angles bluntly rounded, lateral rim denticulate, most conspicuous anteriorly; disc slightly convex; sagittal, anterior and posterior portions densely punctate; remaining disc portions moderately densely punctate; interstices glabrous; anterior and posterior sublateral foveae slightly impressed, rather inconspicuous; entire lateral portions slightly deflexed, rugulously bipunctate, partly microstriate.
Elytra elongate, almost parallel-sided apical 0.15–0.70; disc slightly vaulted, sublaterally more abruptly declivitous; elytral margin moderately explanate up to ca. apical 0.15. Elytra with six regularly arranged, not or slightly impressed rows of puncture striae between suture and disc declivity (approx. at the middle of shoulder) and ca. six additional, less regular puncture striae between disc declivity and elytral margin; punctures moderately large and moderately deeply impressed on anterior disc, gradually slightly decreasing in size and degree of impression towards apex and margin; intervals and interstices flat and glabrous; intervals smaller than puncture diameter anteriorly, larger in posterior and lateral portions; apical sutural teeth present or absent, apices separately rounded, sexually dimorphic (Fig.

Ventral side as in Fig.
Male terminal sternite subsemicircular, 0.18 mm wide, not distinguishable from H. saga and similar species (comp.
Aedeagus (Fig.
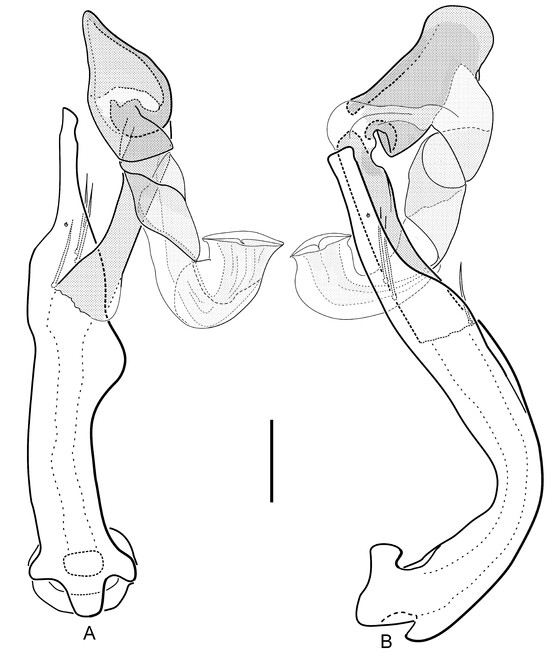
Female tergite X (Fig.
Gonocoxite (Fig.
Spermatheca not examined.
Secondary sexual characters: Female elytral apices produced and separately gently rounded, not acuminate. All femora of male slightly more inflated. Male ventrite VI enlarged (Fig.
Differential Diagnosis
Hydraena dinarica, sp. n. is morphologically very similar to species that are referred to as H. saga complex (sensu
In comparison with all species mentioned above, Hydraena dinarica, sp. n. is unique in the tuba-like 180° bent hyaline distal tube of the aedeagus (Fig.
Within this complex, H. dinarica, sp. n. seems morphologically most similar to the Italian species H. kahleni and H. larissae, especially based on their moderately large contorted aedeagal distal lobe, as well as H. saga on the external habitus. While H. dinarica, sp. n. is slightly larger (2.25–2.45 mm long) than the latter three species (1.95–2.30 mm long), the elytral disc appears slightly flatter and the elytral margin very slightly more explanate in H. dinarica, sp. n. The elytral apices are similar and within the observed variation range in the former species in both sexes. The new species also resembles H. samnitica of almost the same size (especially in the moderately large contorted aedeagal distal lobe), but it is externally distinguishable from H. samnitica by the explanate elytral margin extending almost up to the apex (vs. reaching apical 0.15; the apical area therefore appears more slender in H. dinarica, sp. n.).
On the other hand, H. dinarica, sp. n. seems genetically closest to H. alpicola, H. saga and the H. gracilis Germar, 1824 complex (as defined by
Males can be distinguished as stated above, while in females of H. dinarica, sp. n., the gonocoxite (Fig.
Externally, the new species also resembles other representatives of the "Haenydra" lineage which might occur sympatrically, but differ in certain characters: Hydraena bosnica Apfelbeck, 1909 (posteriorly sinuate pronotum, oval elytra;
Hydraena dinarica, sp. n. varies by 0.6% genetic distance (657 bp CO1 barcode from the most similar congeners H. alpicola and H. saga and by 3.3% from the morphologically similar H. larissae (Suppl. material
Habitat
This species was collected from a mountain stream in a forested, undisturbed karst area at an altitude of about 1220 m a.s.l. (Fig.
Distribution
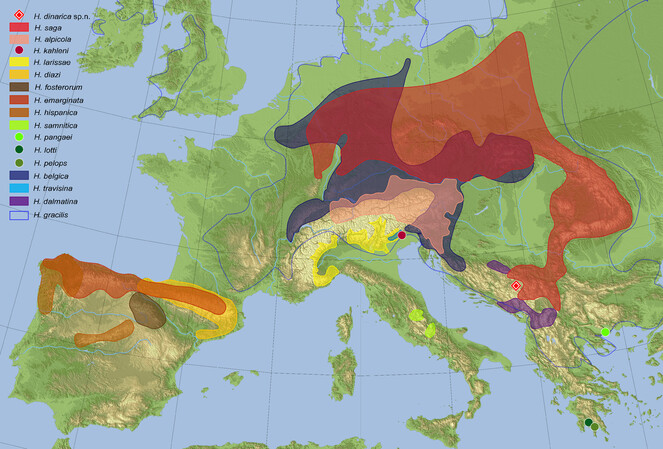
So far only known from the type locality at the northern slopes of Durmitor Mt., Montenegro (Fig.
Etymology
The species is named after the Dinaric Alps, or Dinarides, a karst mountain range where Durmitor Mt. and the type locality of the new species are situated. The epithet is used as an adjective meaning "of the Dinaric Alps".
Analysis
DNA barcoding
The amplification of all sequences but one (H67), with LCO1490 & HC02198 primers and the applied protocols, was successful (Table
GenBank accession numbers of Hydraena COI mtDNA barcode sequences generated in this study.
| Species | Specimen | Site | Voucher | Sequencing | ENA | BOLD |
| H. dinarica sp. n. | female | MNE18c | H68 | Run1 BC12 | MT784148 | TXEX049-20 |
| H. dinarica sp. n. | male | MNE18c | H67 | - | - | - |
| H. subintegra | male | MNE17c | H63 | Run1 BC09 | MT784149 | TXEX051-20 |
| H. nigrita | male | MNE17b | H61 | Run1 BC08 | MT784150 | TXEX052-20 |
| H. sp. (cf. britteni) | female | MNE10l | H65 | Run1 BC10 | MT784151 | TXEX053-20 |
| H. sp. | female | MNE13k | H66 | Run1 BC11 | MT784152 | TXEX054-20 |
| H. morio | male | MNE17b | H62 | Run2 BC08 | MT784154 | TXEX055-20 |
| H. morio | female | MNE17b | H64 | Run2 BC09 | MT784155 | TXEX056-20 |
| H. morio | male | MNE17b | H71 | Sanger | MT784156 | TXEX057-20 |
| H. morio | male | MNE17b | H70 | Sanger | MT784157 | TXEX058-20 |
| H. biltoni | male | MNE21c | H69 | Sanger | MT784158 | TXEX050-20 |
| H. minutissima | male | MNE17b | H60 | Run1 BC07 | MT784159 | TXEX059-20 |
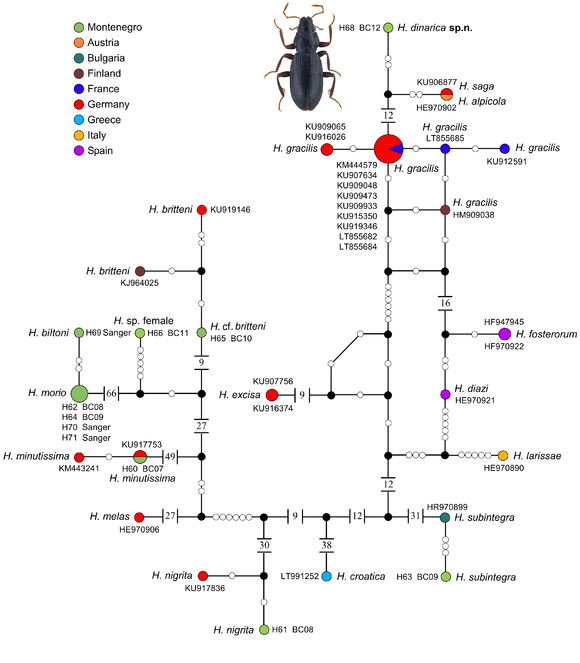
One repository sequence (KU906877), labelled "Hydraena gracilis", does not cluster together with the remaining samples of this species, but rather with H. alpicola Pretner, 1931 (a species found around the northern and eastern Alps and first recognised by
Interspecific genetic distances ranged from 0.0–17.8% (Suppl. material
Discussion
Due to the recent speciation, it is not surprising that we can barely delineate the species complex nor the individual new species by employing a 2% or 3% species delineation threshold of the mtDNA barcode as originally proposed (
Mitochondrial genes, in general and thus standard barcodes, often lag behind in terms of lineage sorting compared to nDNA involved in speciation (e.g.
The habitat of Hydraena dinarica is quite typical for some highly specialized representatives of the "Haenydra" lineage. A similar habitat was also figured for the recently discovered Hydraena kucinici Mičetić Stanković & Jäch in the Republic of Macedonia (
Hydraena dinarica clearly belongs to the "Haenydra" lineage (formerly regarded as subgenus by
Based on the DNA data available (e.g. current study,
Many of these species are known to be highly endemic and all of them are young species, some of the closest relatives, such as H. saga and H. alpicola, have just split recently during the Pleistocene (about 50,000 years ago, based on data of
Its rarity, its presumably very limited distribution range and its special habitat association suggest that the new species is particularly vulnerable to climate change and habitat destruction.
Acknowledgements
Specimens were collected under Permit 02-UPI-662/4 kindly issued by the Environment Protection Agency of Montenegro. The first author wants to thank the curators Dr. Manfred A. Jäch (Natural History Museum Vienna) and Olaf Jäger (Senckenberg Natural History Collections Dresden) for access to and loan of their collections. The whole team would like to thank the staff of Etno selo Šljeme for their hospitality and for allowing us to set up a laboratory in a part of their restaurant.
Author contributions
Conceptualisation, HF, MD & IN; collection and identification of material, HF, RdV, CGT, HL, RL, MFF, MG & CVP; formal species description, HF & RdV; MinION sequencing, MP, SM, MD; preparation of Sanger sequencing, HF, ED, MRS; molecular data analysis, ED & MRS; writing—original draft preparation, HF, RdV, CGT, MP, SM, IN; writing—review and editing, all authors.
References
- Using phylogenies to trace the geographical signal of diversification.Journal of Biogeography44(10):2236‑2246. https://doi.org/10.1111/jbi.13035
- Contributiones ad Coleopterorum faunam peninsulae balcanicae.Sarajevo. Glasnik Zemaljskog Muzeja Bosne i Hercegovine12:642‑664.
- Suggestions for a molecular biodiversity assessment of South East Asian freshwater invertebrates: Lessons from the megadiverse beetles (Coleoptera).Journal of Limnology72(S2):866‑880. https://doi.org/10.4081/jlimnol.2013.s2.e4
- Remarks on the genus Hydraena and revision of the subgenus Phothydraena (Coleoptera: Hydraenidae).Annales de Limnologie22:181‑193. https://doi.org/10.1051/limn/1986016
- Hydraena lotti sp. nov., a new member of the Haenydra lineage from the Peloponnese (Greece), with additional records of Hydraena species in the region (Coleoptera, Hydraenidae).Zootaxa3637(1):29‑38. https://doi.org/10.11646/zootaxa.3637.1.3
- TCS: A computer program to estimate gene genealogies.Molecular Ecology9:1657‑1659. https://doi.org/10.1214/aos/1013699998
- Reconstruction of the pleistocene glaciers of Mt. Durmitor in Montenegro.Acta Geographica Slovenica49(2):263‑289. https://doi.org/10.3986/AGS49202
- Due nuove specie paleartiche appartenenti al genere Haenydra Rey. XXV Contributo alla conoscenza degli Hydraenidae.Bulletin & Annales de la Société Royale Belge d'Entomologie127:217‑221.
- Ancora sui caratteri sessuali secondari di alcuni Coleotteri.Rivista Coleotterologica Italiana2(12):233‑254.
- Hydraena (Hydraenopsis) ateneo, new species (Coleoptera, Hydraenidae) and other aquatic polyphaga from a small habitat patch in a highly urbanized landscape of Metro Manila, Philippines.ZooKeys329:9‑21. https://doi.org/10.3897/zookeys.329.5955
- Clues to aquatic beetle research in SE Asia: A multitude of ecological niches, microhabitats and deduced field sampling techniques.Entomology, Ornithology & Herpetology4(4):166. https://doi.org/10.4172/2161-0983.1000166
- Three new species of Grouvellinus Champion, 1923 from Maliau Basin, Sabah, Borneo, discovered by citizen scientists during the first Taxon Expedition (Insecta, Coleoptera, Elmidae.ZooKeys754:1‑21. https://doi.org/10.3897/zookeys.754.24276
- Beiträge zur Kenntniss der paläarktischen Hydrophiliden.Verhandlungen der Zoologisch - Botanischen Gesellschaft in Wien51:312‑332.
- Insectorum species novae aut minus cognitae, descriptionibus illustratae. Coleoptera.J.C. Hendelii et filii, Halae0‑624.
- BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT.Nucleic Acids Symposium Series4:95‑98.
- Biological identifications through DNA barcodes.Proceedings of the Royal Society B270:313‑321. https://doi.org/10.1098/rspb.2002.2218
- Mitochondrial cox1 sequence data reliably uncover patterns of insect diversity but suffer from high lineage-idiosyncratic error rates.PLOS One5(12). https://doi.org/10.1371/journal.pone.0014448
- A comprehensive DNA barcode database for Central European beetles with a focus on Germany: adding more than 3500 identified species to BOLD.Molecular Ecology Resources15:795‑818. https://doi.org/10.1111/1755-0998.12354
- The Hydraena (Haenydra) gracilis Germar species complex (Insecta: Coleoptera:Hydraenidae).Annalen des Naturhistorischen Museums in Wien97B:177‑190. URL: https://www.zobodat.at/pdf/ANNA_97B_0177-0190.pdf
- Beschreibung neuer Hydraena-Arten aus Jugoslawien (Coleoptera, Hydraenidae).Nachrichtenblatt der Bayerischen Entomologen35(3):65‑69.
- New and little known Palearctic species of the genus Hydraena (s. l.) Kugelann (Coleoptera: Hydraenidae).Koleopterologische Rundschau62:77‑125.
- New Hydraenidae from the Peloponnese (Greece) (Coleoptera: Hydraenidae).Entomological Problems26(1):39‑42.
- New and little known Palearctic species of the genus Hydraena VI (Coleoptera: Hydraenidae).Entomological Problems31(2):155‑162.
- 11.1. Hydraenidae Mulsant, 1844. In:Handbook of Zoology, Volume IV (Part 38), Coleoptera, Beetles, Volume 1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, polyphaga partim).Walter de Gruyter,Berlin, New York,471-496pp. https://doi.org/10.1515/9783110904550
- New and little known Palearctic species of the genus Hydraena (s.l.) Kugelann X. Descriptions of four new species from southern Europe, and taxonomic review of the H. subintegra species complex (Coleoptera: Hydraenidae).Koleopterologische Rundschau82:95‑113.
- Hydraenidae. In:Catalogue of Palaearctic Coleoptera. Vol. 2/1. Hydrophiloidea – Staphylinoidea. Revised and updated edition.Brill,Leiden,130-162pp.
- New and little known Palearctic species of the genus Hydraena (s.l.) Kugelann, 1794 XII. Description of a new species of the H. saga complex from Italy (Coleoptera: Hydraenidae).Koleopterologische Rundschau87:37‑50.
- Hydraena britteni, sp. nov., a new British beetle.The Entomologist's Monthly Magazine43:79‑81.
- Monographische Revision der Gattung Hydraena.Linnaea Entomologica4:156‑190.
- Verzeichniß der in einigen Gegenden Preußens bis jetzt entdeckten Käfer-Arten, nebst kurzen Nachrichten von denselben.Neuestes Magazin für die Liebhaber der Entomologie1:513‑582.
- MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms.Molecular Biology and Evolution35(6):1547‑1549. https://doi.org/10.1093/molbev/msy096
- PopART: Full-feature software for haplotype network construction.Methods in Ecology and Evolution6(9):1110‑1116. https://doi.org/10.1111/2041-210X.12410
- A rapid and accurate MinION-based workflow for tracking species biodiversity in the field.Genes10(6):468. https://doi.org/10.3390/genes10060468
- DNA barcoding: error rates based on comprehensive sampling.PLOS Biology3(12):e422. https://doi.org/10.1371/Journal.Pbio.0030422
- Description of a new species of the "Haenydra" lineage (Coleoptera: Hydraenidae) from the Republic of Macedonia.Zootaxa3503:82‑88. https://doi.org/10.11646/zootaxa.3503.1.6
- Beyond barcodes: Complex DNA taxonomy of a South Pacific Island radiation.Proceedings of the Royal Society B: Biological Sciences273:887‑893. https://doi.org/10.1098/rspb.2005.3391
- Nouvelles notes sur quelques Hydraena paléarctiques.Bulletin & Annales de la Société Royale Belge d'Entomologie70:218‑228.
- Notes sur quelques Hydraena paléarctiques.Bulletin & Annales de la Société Royale Belge d'Entomologie69:367‑386.
- Notes sur quelques Hydraena circum–méditerranéennes.Bulletin & Annales de la Société Royale Belge d'Entomologie71:63‑73.
- Barcoding beetles: a regional survey of 1872 species reveals high identification success and unusually deep interspecific divergences.PLOS One9(9):e108651. https://doi.org/10.1371/journal.pone.0108651
- Life on the effective bubble: exocrine secretion delivery systems (ESDS) and the evolution and classification of beetles in the family Hydraenidae (Insecta: Coleoptera).Annals of the Carnegie Museum66:89‑207.
- Über die angebliche Variabilität der Hydraena gracilis Germar.Coleopterologisches Centralblatt5(2):81‑106.
- Description de Coléoptères nouveaux ou peu connus de la tribu des Palpicornes.Annales de la Société Linnéenne de Lyon et des Sociétés botaniques de Lyon31(1884):13‑32.
- The geography of speciation in narrow range endemics of the Haenydra lineage (Coleoptera, Hydraenidae, Hydraena).Journal of Biogeography38:502‑516. https://doi.org/10.1111/j.1365-2699.2010.02417.x
- Using taxonomic consistency with semiautomated data preprocessing for high quality DNA barcodes.Methods in Ecology and Evolution8:1878‑1887. https://doi.org/10.1111/2041-210X.12824
- Three new minute leaf litter beetles discovered by citizen scientists in Maliau Basin, Malaysian Borneo (Coleoptera: Leiodidae, Chrysomelidae).Biodiversity Data Journal5:e21947. https://doi.org/10.3897/BDJ.5.e21947
- Illustrations of British entomology; or, a synopsis of indigeneous insects: containing their generic and specific distributions; with an account of their metamorphoses, times of appearance, localities, food, and economy, as far as practicable. In:Mandibulata 2.Baldwin and Craddock,London,87pp. https://doi.org/10.5962/bhl.title.8133
- Molecular and morphological analyses reveal two new species of the Hydraena saga–emarginata complex (Coleoptera Hydraenidae, Hydraeninae) from Spain and France.Zootaxa2760:29‑38. https://doi.org/10.11646/zootaxa.2760.1.3
- Molecular phylogeny and diversification of the "Haenydra" lineage (Hydraenidae, genus Hydraena), a north-Mediterranean endemic-rich group of rheophilic Coleoptera.Molecular Phylogenetics and Evolution61:772‑783. https://doi.org/10.1016/j.ympev.2011.07.020
- A revision of Hydraena species of the "Haenydra" lineage (Coleoptera, Hydraenidae).Zootaxa3607(1):1‑173. https://doi.org/10.11646/zootaxa.3607.1.1
- A molecular phylogeny of the cosmopolitan hyperdiverse genus Hydraena Kugelann (Coleoptera, Hydraenidae).Systematic Entomology38:192‑208. https://doi.org/10.1111/j.1365-3113.2012.00654.x
Supplementary materials
Specimens sequenced in the field by the use of the ONTrack MinION pipeline are indicated by their "H[number]" code. Sequences of Hendrich et al. (2015), Pentinsaari et al. (2014), Ribera (2011), Rulik et al. (2017), Trizzino et al. (2011b; 2013b), are indicated by their respective GenBank accession numbers.