|
Biodiversity Data Journal :
Taxonomic Paper
|
|
Corresponding author:
Academic editor: Edward Baker
Received: 14 Jan 2016 | Accepted: 29 Feb 2016 | Published: 07 Mar 2016
© 2016 Ming Kai Tan, Khairul Kamaruddin
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Tan M, Kamaruddin K (2016) Redescription of the little-known grasshopper Willemsella (Acrididae, Hemiacridinae) from Peninsular Malaysia. Biodiversity Data Journal 4: e7775. https://doi.org/10.3897/BDJ.4.e7775
|
|
Abstract
Background
Catantopinae is a huge subfamily and is in need of a major revision. We contribute to the taxonomy of the subfamily by reviewing one poorly known genus, Willemsella Miller, 1934. This is a monotypic genus so far found only in Peninsular Malaysia.
New information
Rediscovery and new locality records of the species was reported here. The species is redescribed in accordance to modern criteria of taxonomy. Male phallic complex is also described for the first time. The subfamily position of the species is also verified and is more correctly placed under Hemiacridinae.
Keywords
Orthoptera, Catantopinae, male phallic complex, new locality record, Southeast Asia
Introduction
Catantopinae is a huge subfamily consisting of around 16 tribes and about 382 genera and is distributed throughout the old world (
Willemsella is a monotypic genus so far and is found only in Peninsular Malaysia. Described in 1934 based on a pair of copulating adults in Dusan Tua, Selangor (
While the original description of the genus and its type species is substantial, description of the male phallic complex is lacking. It was also not examined in the major revision of phallic complexes of grasshoppers (
Materials and methods
New material was collected by the authors in the highlands of Peninsular Malaysia: Bukit Fraser (Pahang) from 2012 to 2013 and Bukit Larut (Perak) in 2015. The grasshoppers were mainly collected by opportunistic collection and sweep netting among tall grasses in the day or night. The specimens were either preserved in absolute analytic alcohol and/ or air dried and pinned. Photographic images were done with a Canon EOS 500D digital SLR camera with a compact-macro lens EF 100mm 1:2.8 USM, then edited using Adobe Photoshop. Images of the male genitalia were done using the Visionary Digital System. Scales are given with the images. Male genitalia were preserved in glycerine. Whenever possible, in-situ images were also taken using a Canon EOS 500D digital SLR camera with a compact-macro lens. Measurements of dried pinned specimen were made using Vernier calipers. In the measurement section, the following terms were used: BL = body length; VW = vertex width between eyes; FRW = frontal ridge width between antennae; PL = pronotum length; PZL = prozona length; MZL = metazona length; PW = pronotum width; TL = tegmen length; HFL = hind femur length; HFW = hind femur width (maximum); HTL = hind tibia length. The specimens are deposited in the Zoological Reference Collection (ZRC), Lee Kong Chian Natural History Museum (former Raffles Museum of Biodiversity Research).
For description and illustration captures, we use the following terms as already used by
Epiphallus:
B Bridge
A Ancora
An Anterior projection
L Lophus
Ectophallus:
Dsv Dorsal ectophallic sclerite
Esv Ventral ectophallic sclerite
Apd Apodeme
Ac Arch of cingulum
Ap Apical valve of penis (= aedeagus)
Bp Basal valve of penis (= endoparameres, endophallic plates)
Taxon treatment
Willemsella bicolor
Nomenclature
The genus should be placed more correctly under Hemiacridinae, rather than Catantopinae. In Willemsella, the basal and apical penis valves are separated, diagnostic for species in Hemiacridinae. In Catantopinae, the basal and penis valves are connected. The genus also exhibits a series of regular transverse veinlets near the radial area of tegmen, similar to those found in Hemiacridinae and usually absent in Catantopinae. Willemsella also shares characters of Catantopinae: presence of prosternal spine, rounded mesosternal lobes and absence of intercalary veins in the medial area of tegmen.
-
scientificName: Willemsella bicolor; namePublishedIn:1934; nameAccordingTo:Miller; kingdom:Animalia; phylum:Arthropod; class:Insecta; order:Orthoptera; family:Acrididae; genus:Willemsella; specificEpithet:bicolor; country:Peninsular Malaysia; stateProvince:Perak; locality:Bukit Larut (Maxwell Hill); verbatimElevation:1056 m; decimalLatitude:4.86184; decimalLongitude:100.79276; samplingProtocol:active collecting; eventDate:06/18/2015; eventTime:1053-1201; individualCount:1; sex:male; lifeStage:adult; catalogNumber:LAR.15.53; recordedBy:Ming Kai Tan et al.; disposition:ZRC; identifiedBy:M. K. Tan; dateIdentified:2015
-
scientificName: Willemsella bicolor; namePublishedIn:1934; nameAccordingTo:Miller; kingdom:Animalia; phylum:Arthropod; class:Insecta; order:Orthoptera; family:Acrididae; genus:Willemsella; specificEpithet:bicolor; country:Peninsular Malaysia; stateProvince:Perak; locality:Bukit Larut (Maxwell Hill); verbatimElevation:920.4 m; decimalLatitude:4.86626; decimalLongitude:100.78865; samplingProtocol:active collecting; eventDate:09/19/2015; eventTime:1925-2049; individualCount:1; sex:male; lifeStage:adult; catalogNumber:LAR.15.189; recordedBy:Ming Kai Tan et al.; disposition:ZRC; identifiedBy:M. K. Tan; dateIdentified:2015
-
scientificName: Willemsella bicolor; namePublishedIn:1934; nameAccordingTo:Miller; kingdom:Animalia; phylum:Arthropod; class:Insecta; order:Orthoptera; family:Acrididae; genus:Willemsella; specificEpithet:bicolor; country:Peninsular Malaysia; stateProvince:Perak; locality:Bukit Larut (Maxwell Hill); verbatimElevation:1001 m; decimalLatitude:4.86289; decimalLongitude:100.79163; samplingProtocol:active collecting; eventDate:09/20/2015; eventTime:1043-1236; individualCount:1; sex:male; lifeStage:adult; catalogNumber:LAR.15.199; recordedBy:Ming Kai Tan et al.; disposition:ZRC; identifiedBy:M. K. Tan; dateIdentified:2015
-
scientificName: Willemsella bicolor; namePublishedIn:1934; nameAccordingTo:Miller; kingdom:Animalia; phylum:Arthropod; class:Insecta; order:Orthoptera; family:Acrididae; genus:Willemsella; specificEpithet:bicolor; country:Peninsular Malaysia; stateProvince:Perak; locality:Bukit Larut (Maxwell Hill); verbatimElevation:1097 m; decimalLatitude:4.86167; decimalLongitude:100.79434; samplingProtocol:active collecting; eventDate:09/20/2015; eventTime:1928-2055; individualCount:1; sex:male; lifeStage:adult; catalogNumber:LAR.15.203; recordedBy:Ming Kai Tan et al.; disposition:ZRC; identifiedBy:M. K. Tan; dateIdentified:2015
-
scientificName: Willemsella bicolor; namePublishedIn:1934; nameAccordingTo:Miller; kingdom:Animalia; phylum:Arthropod; class:Insecta; order:Orthoptera; family:Acrididae; genus:Willemsella; specificEpithet:bicolor; country:Peninsular Malaysia; stateProvince:Perak; locality:Bukit Larut (Maxwell Hill); verbatimElevation:1097 m; decimalLatitude:4.86167; decimalLongitude:100.79434; samplingProtocol:active collecting; eventDate:09/20/2015; eventTime:1928-2055; individualCount:1; sex:female; lifeStage:adult; catalogNumber:LAR.15.210; recordedBy:Ming Kai Tan et al.; disposition:ZRC; identifiedBy:M. K. Tan; dateIdentified:2015
-
scientificName: Willemsella bicolor; namePublishedIn:1934; nameAccordingTo:Miller; kingdom:Animalia; phylum:Arthropod; class:Insecta; order:Orthoptera; family:Acrididae; genus:Willemsella; specificEpithet:bicolor; country:Peninsular Malaysia; stateProvince:Perak; locality:Bukit Larut (Maxwell Hill); verbatimElevation:1052 m; decimalLatitude:4.86191; decimalLongitude:100.79305; samplingProtocol:active collecting; eventDate:09/20/2015; eventTime:2055-2136; individualCount:1; sex:female; lifeStage:adult; catalogNumber:LAR.15.211; recordedBy:Ming Kai Tan et al.; disposition:ZRC; identifiedBy:M. K. Tan; dateIdentified:2015
-
scientificName: Willemsella bicolor; namePublishedIn:1934; nameAccordingTo:Miller; kingdom:Animalia; phylum:Arthropod; class:Insecta; order:Orthoptera; family:Acrididae; genus:Willemsella; specificEpithet:bicolor; country:Peninsular Malaysia; stateProvince:Pahang; locality:Bukit Fraser; verbatimElevation:1214 m; decimalLatitude:3.71944; decimalLongitude:101.72936; samplingProtocol:active collecting; eventDate:12/27/2012; eventTime:1338; individualCount:1; sex:male; lifeStage:adult; catalogNumber:FRA.12.88; recordedBy:Ming Kai Tan et al.; disposition:ZRC; identifiedBy:M. K. Tan; dateIdentified:2015
-
scientificName: Willemsella bicolor; namePublishedIn:1934; nameAccordingTo:Miller; kingdom:Animalia; phylum:Arthropod; class:Insecta; order:Orthoptera; family:Acrididae; genus:Willemsella; specificEpithet:bicolor; country:Peninsular Malaysia; stateProvince:Pahang; locality:Bukit Fraser; verbatimElevation:1203 m; decimalLatitude:3.71952; decimalLongitude:101.72937; samplingProtocol:active collecting; eventDate:12/27/2012; eventTime:1346; individualCount:1; sex:male; lifeStage:adult; catalogNumber:FRA.12.90; recordedBy:Ming Kai Tan et al.; disposition:ZRC; identifiedBy:M. K. Tan; dateIdentified:2015
-
scientificName: Willemsella bicolor; namePublishedIn:1934; nameAccordingTo:Miller; kingdom:Animalia; phylum:Arthropod; class:Insecta; order:Orthoptera; family:Acrididae; genus:Willemsella; specificEpithet:bicolor; country:Peninsular Malaysia; stateProvince:Pahang; locality:Bukit Fraser; verbatimElevation:1209 m; decimalLatitude:3.71943; decimalLongitude:101.72935; samplingProtocol:active collecting; eventDate:12/29/2012; eventTime:2005; individualCount:1; sex:male; lifeStage:adult; catalogNumber:FRA.12.112; recordedBy:Ming Kai Tan et al.; disposition:ZRC; identifiedBy:M. K. Tan; dateIdentified:2015
-
scientificName: Willemsella bicolor; namePublishedIn:1934; nameAccordingTo:Miller; kingdom:Animalia; phylum:Arthropod; class:Insecta; order:Orthoptera; family:Acrididae; genus:Willemsella; specificEpithet:bicolor; country:Peninsular Malaysia; stateProvince:Pahang; locality:Bukit Fraser; verbatimElevation:1227 m; decimalLatitude:3.71836; decimalLongitude:101.72987; samplingProtocol:active collecting; eventDate:05/19/2013; eventTime:1409; individualCount:1; sex:female; lifeStage:adult; catalogNumber:FRA.13.35; recordedBy:Ming Kai Tan et al.; disposition:ZRC; identifiedBy:M. K. Tan; dateIdentified:2015
Description
Adult habitus as shown in Fig.










Male. Epiproct broadly tongue-shaped with an apical lobe; median sulcus shallow, broad and feeble, only in the basal half bordered by broad parallel carinae; apical lobe triangular with obtuse apex (Fig.

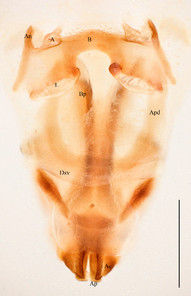
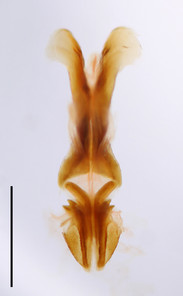



Willemsella bicolor Miller, 1934. Scale bars: 1 mm.
b: Male phallic complex in dorsal view.
c: Basal and apical penis valves in dorsal view.
d: Female abdominal apex in dorsal view.
e: Female abdominal apex in lateral view.
f: Female abdominal apex in ventral view.
Female. Tenth abdominal tergite with posterior margin rounded-triangularly excised, without lateral lobe (Fig.
Colouration (Fig.
Measurement of alcohol-preserved dry-pinned specimens, in mm (mean in bracket). Male (n=3) BL: 18.4-19.9 (19.1), VW: 0.5-0.7 (0.6), FRW: 0.1-0.2 (0.1), PL: 3.4-4.2 (3.8), PZL: 2.3-2.6 (2.4), MZL: 1.1-1.6 (1.4), PW: 3.0-3.4 (3.2), TL: 7.9-8.7 (8.4), HFL: 12.7-13.6 (13.1), HFW: 2.7-2.9 (2.8), HTL: 11.5-12.5 (12.0); female (n=2) BL: 21.9-22.4 (22.2), VW: 1.0, FRW: 0.5, PL: 5.2-5.5 (5.4), PZL: 3.4-3.9 (3.7), MZL: 1.6-1.8 (1.7), PW: 4.1-4.6 (4.4), TL: 9.9-10.2 (10.1), HFL: 15.4-15.9 (15.7), HFW: 3.2-3.4 (3.3), HTL: 14.0-14.2 (14.1).
Diagnosis
Prosternal spine distinct, flattened and triangular basally, narrowing into a long and subacute apex. Radial area of tegmen with a series of regular transverse veinlets. Anchora (A) articulated, extending in less than 90º-angle to epiphallus bridge, apex obtuse from dorsal view; membrane between lophi sparsely granulated. Anterior projection (An) of epiphallus swollen and somewhat sparsely granular, forming at less than 90º-angle to the anchora. Basal and apical penis valves separated. Antennal segments bright red. Lateral lobes with a basal bright yellow spot extending from the anterior to the posterior margins of lateral lobe.
Distribution
Malay Peninsula: Bukit Larut, Perak; Bukit Fraser, Pahang; Dusan Tua, Selangor (
Ecology
Little is known about the ecology of this grasshopper species. they are fairly common among tall grasses in Bukit Fraser and Bukit Larut, suggesting that they may prefer high elevations.
Conservation
So far endemic to Peninsular Malaysia.
Acknowledgements
The authors thank Siew Tin Toh and Illias bin Abdul Rani for assistance in field collection; LKCNHM for access to the ZRC to examine specimens and the Cryogenic Laboratory to use the VD System. Permission for the collection of material was granted by the Research Promotion and Co-Ordination Committee, Economic Planning Unit, Prime Minister’s Department (UPE: 40/200/19/3103 and 40/200/19/2923) and supported by the Institute for Biodiversity, Department of Wildlife and National Parks (Perhilitan). The work of MKT was partly supported by Lady Yuen Peng McNeice Graduate Fellowship.
References
- The phallic complex in Acridoidea (Orthoptera) in relation to taxonomy.Transactions of the Royal entomological Society of London108(7):223‑270. https://doi.org/10.1111/j.1365-2311.1956.tb02270.x
- A preliminary revision of the families and subfamilies of Acridoidea (Orthoptera, Insecta).Bulletin of the British Museum (Natural History)10(9):27‑419. https://doi.org/10.5962/bhl.part.16264
- Classification of the acridomorphoid insects.Faringdon, Oxon (E. W. Classey,171pp.
- Orthoptera species file online. Version 5 (5.0). http://orthoptera.speciesfile.org/HomePage/Orthoptera/HomePage.aspx
- Notes on Malayan Acrididae and description of some new genera and species.Journal of the Federation of Malay State Museum17:526.
- Orthoptera of Fraser’s Hill, Peninsular Malaysia.Lee Kong Chian Natural History Museum, National University of Singapore,Singapore,88pp.