|
Biodiversity Data Journal :
Species inventory
|
The Influence of Landscape Heterogeneity - Ground Beetles (Coleoptera: Carabidae) in Fthiotida, Central Greece
|
Corresponding author:
Academic editor: Stephen Venn
Received: 26 Mar 2014 | Accepted: 10 Apr 2014 | Published: 11 Apr 2014
© 2014 Anna Chapman
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Chapman A (2014) The Influence of Landscape Heterogeneity - Ground Beetles (Coleoptera: Carabidae) in Fthiotida, Central Greece. Biodiversity Data Journal 2: e1082. https://doi.org/10.3897/BDJ.2.e1082
|
|
Abstract
Pitfall traps were used to sample Carabidae in agricultural land of the Spercheios valley, Fthiotida, Central Greece. Four pairs of cultivated fields were sampled. One field of each pair was located in a heterogeneous area and the other in a more homogeneous area. Heterogeneous areas were composed of small fields. They had high percentages of non-cropped habitats and a high diversity of land use types. Homogeneous areas were composed of larger fields. They had lower percentages of non-cropped habitats and a lower diversity of land use types. One pair of fields had been planted with cotton, one with maize, one with olives and one with wheat. Altogether 28 carabid species were recorded. This paper describes the study areas, the sampling methods used and presents the data collected during the study. Neither heterogeneous nor homogeneous areas had consistently higher abundance levels, activity density levels, species richness levels or diversity levels. However, significant differences were seen in some of the comparisons between heterogeneous and homogeneous areas.
Keywords
Carabidae, Coleoptera, agriculture, farmland, cotton, maize, olives, wheat, cultivation, crops, heterogeneity, landscape, Greece
Introduction
The level of heterogeneity in agricultural landscapes can influence farmland wildlife (
Locally, soil moisture and soil type have a large impact on carabid distributions (
Small fields are thought to be easier for carabids to recolonize after disturbance, due to the shorter dispersal distances involved. Additionally, landscapes with small fields are likely to have high levels of land use diversity, which will create refuges for carabids in times of disturbance. This is because cultivation practices take place at different times in different crops, so a diverse landscape will always have some undisturbed habitat, while cultivation practices take place elsewhere (
Another aspect of landscape heterogeneity that is important for carabids, is the presence of non-cropped habitat. This may take the form of grassy field margins, hedgerows and areas of semi-natural habitat, such as fallow, woodland and wasteland. Although carabids live in the crops during the vegetation season, they are known to use field margins to hibernate in (
In Greece, agriculture is often extensive and small in scale, with large-scale, intensive farming occurring only on the flatter and more fertile land. This means that the country has relatively high levels of habitat richness (
This study aims to identify closely matched and situated areas of heterogeneous and homogeneous farmland. These areas will then be compared using landscape analysis, so that the different aspects of their heterogeneity can be examined. Finally, matched fields of the same crop types within the heterogeneous and homogeneous areas will be compared, to see how they differ in terms of their carabid abundance, activity density, species richness and diversity. In this way, it may be determined whether high levels of landscape heterogeneity benefit Carabidae at a local level, within individual fields.
Materials and methods
Study Areas and Sampled Fields
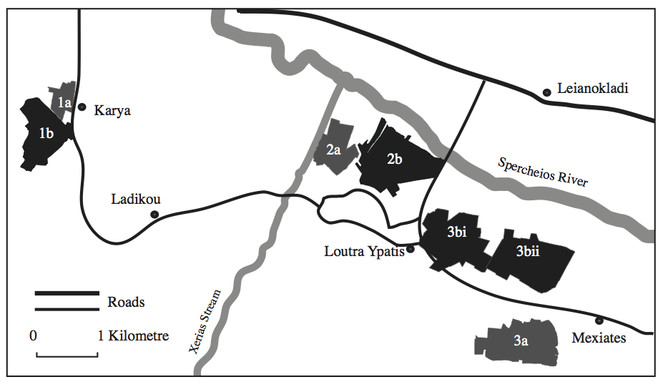
The study was conducted on agricultural land in the Spercheios valley, Fthiotida, Central Greece. Four pairs of fields were sampled for Carabidae between May and October in 2007. One pair of the sampled fields had been planted with cotton, one with maize, one with olives and one with wheat.
One field of each pair was located in a heterogeneous area, which had small field sizes, a large amount of non-cropped habitat and a high level of land use diversity. The other field of each pair was located in a more homogeneous area, which had larger field sizes, less non-cropped habitat and a lower level of land use diversity (Suppl. material
The heterogeneous and homogeneous areas were matched, in order to be as similar as possible regarding all factors apart from their heterogeneity. They were matched according to their mean elevation, their distances to villages and roads, as well as to large expanses of woodlandand wasteland.
Fig.
The sampled fields chosen from within the heterogeneous and homogeneous areas were also matched regarding their crop type, soil type, previous crop type, agrochemical treatment, harvesting time, elevation and whether or not they were irrigated. The data used to match the fields, along with the size and location of each field, are provided in Table
The size and location of each of the sampled fields. Also the data used to match fields in heterogeneous and homogeneous areas.
| Sampled Field - (Study Area) | Field Size (ha) | Location - (Trap Line Coordinates) | Mean Elevation of Field (m) | Dominant Soil Type | Insecticide | Fertilizer | Previous Crop | Harvest Time | Irrigation |
|---|---|---|---|---|---|---|---|---|---|
| Cotton a - (3a) | 0.72 | 38°52'51.84"N 22°17'50.77"E | 51 | Poorly sorted, very coarse sand | Phosalone | 11-15-15 | Cotton | Late October | Yes |
| Cotton b - (3bii) | 2.16 | 38°53'35.73"N 22°17'46.68"E | 36 | Poorly sorted, very coarse sand | Phosalone | 11-15-15 | Cotton | Late October | Yes |
| Maize a - (1a) | 0.08 | 38°54'57.81"N 22°12'35.62"E 38°54'58.13"N 22°12'35.83"E | 97 | Very coarse, silty, very coarse sand | None | 10-20-10, Lime | Maize | Mid September | Yes |
| Maize b - (1b) | 4.76 | 38°54'39.10"N 22°12'24.85"E | 105 | Very coarse, silty, very coarse sand - Very coarse, silty coarse sand. | None | 23-8-6, 0.5 Zn | Maize | Mid September | Yes |
| Olives a - (2a) | 0.14 | 38°54'34.25"N 22°15'38.14"E 38°54'34.31"N 22°15'37.61"E | 80 | Poorly sorted, very coarse sand | None | None | Olives | Late November | No |
| Olives b - (2b) | 10.37 | 38°54'27.91"N 22°16'12.65"E | 70 | Poorly sorted, very coarse sand - Poorly sorted, medium sand | None | None | Olives | Late November | No |
| Wheat a - (3a) | 0.36 | 38°52'51.67"N 22°17'44.45"E | 52 | Poorly sorted, medium sand | None | None | Alfalfa | Early June | No |
| Wheat b - (3bi) | 1.81 | 38°53'46.91"N 22°16'57.99"E | 53 | Poorly sorted, coarse sand | None | None | Alfalfa | Early June | No |
The land use maps in Figs
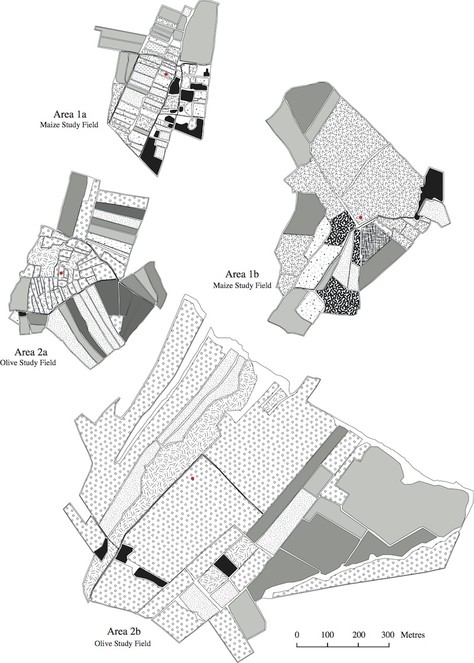
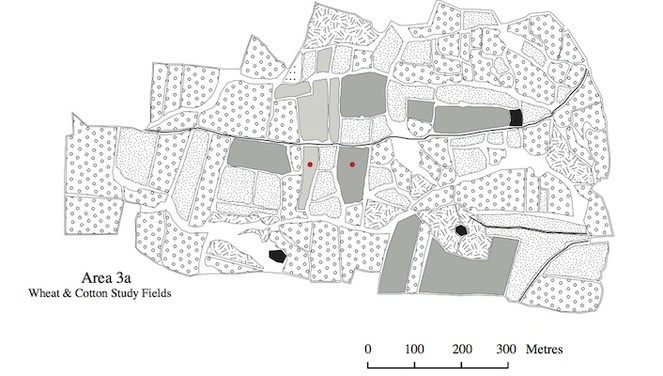
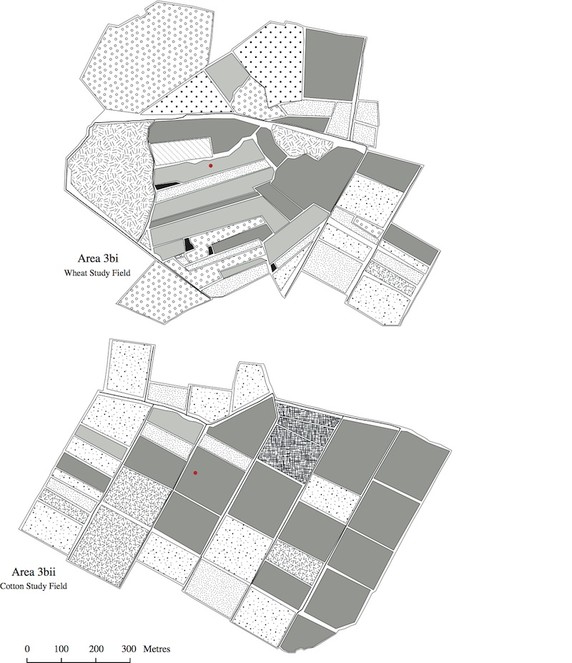
The program FRAGSTATS (
The metric "land use aggregation" which concerns land use configuration, is dimensionless, so therefore lacks units. The same is true for the metric "land use similarity", which indicates how similar the sampled fields were, regarding land use, to other fields in the surrounding areas (
Sampling Procedure
The locations of the pitfall traps within the sampled fields are listed as geographic coordinates in Table
Not all of the pairs of fields were sampled during every 15 day sampling period. This was because access to the fields depended on the cultivation practices taking place at the time. Irrigation, spraying, harvesting, ploughing, pruning and fertilizer application would all prevent access to the fields and the setting of traps. Initially, a pair of alfalfa fields was also sampled, but the frequency of harvesting meant that most of these traps were destroyed before they could be collected. So sampling in these fields was not continued.
If one field of a pair was inaccessible for a given 15 day period, then the other field of that pair was not sampled either. This insured that comparisons between heterogeneous and homogeneous areas were fair, as the same amount of data was obtained in both fields, at the same time of year. In all, 440 traps were set and 391 were recovered successfully. Successful traps were those that were not flooded by irrigation, destroyed by other farming practices or dug up by animals. The dates covered by the 15 day sampling periods, as well as the numbers of traps set and successfully recovered are shown in Table
| Sampled Field - (Study Area) | 15 day Sampling Period | Number of Traps Set | Number of Successful Traps | Number of Traps Used in the Data Analysis |
| Cotton a - (3a) | 22nd May to 6th June | 10 | 10 | 10 |
| 7th June to 22nd June | 10 | 9 | 9 | |
| 9th July to 24th July | 10 | 6 | 5 | |
| 8th Sept to 23rd Sept | 10 | 10 | 10 | |
| 23rd Sept to 8th Oct | 10 | 7 | 6 | |
| Total = 5 periods of 15 days | Total = 50 | Total = 42 | Total = 40 | |
| Cotton b - (3bii) | 22nd May to 6th June | 10 | 10 | 10 |
| 7th June to 22nd June | 10 | 10 | 9 | |
| 9th July to 24th July | 10 | 5 | 5 | |
| 8th Sept to 23rd Sept | 10 | 10 | 10 | |
| 23rd Sept to 8th Oct | 10 | 10 | 6 | |
| Total = 5 periods of 15 days | Total = 50 | Total = 45 | Total = 40 | |
| Maize a - (1a) | 7th June to 22nd June | 10 | 10 | 10 |
| 23rd June to 8th July | 10 | 10 | 9 | |
| 9th July to 24th July | 10 | 10 | 9 | |
| 9th Aug to 24th Aug | 10 | 9 | 8 | |
| 8th Sept - 23rd Sept | 10 | 4 | 4 | |
| Total = 5 periods of 15 days | Total = 50 | Total = 43 | Total = 40 | |
| Maize b - (1b) | 7th June to 22nd June | 10 | 10 | 10 |
| 23rd June to 8th July | 10 | 10 | 9 | |
| 9th July to 24th July | 10 | 9 | 9 | |
| 9th Aug to 24th Aug | 10 | 9 | 8 | |
| 8th Sept to 23rd Sept | 10 | 7 | 4 | |
| Total = 5 periods of 15 days | Total = 50 | Total = 45 | Total = 40 | |
| Olives a - (2a) | 5th May to 20th May | 10 | 8 | 7 |
| 22nd May to 6th June | 10 | 10 | 9 | |
| 23rd June to 8th July | 10 | 10 | 9 | |
| 9th July to 24th July | 10 | 10 | 9 | |
| 25th July to 9th Aug | 10 | 10 | 6 | |
| 8th Oct to 23rd Oct | 10 | 10 | 0 | |
| Total = 6 periods of 15 days | Total = 60 | Total = 58 | Total = 40 | |
| Olives b - (2b) | 5th May to 20th May | 10 | 10 | 7 |
| 22nd May to 6th June | 10 | 10 | 9 | |
| 23rd June to 8th July | 10 | 10 | 9 | |
| 9th July to 24th July | 10 | 10 | 9 | |
| 25th July to 9th Aug | 10 | 10 | 6 | |
| 8th Oct to 23rd Oct | 10 | 9 | 0 | |
| Total = 6 periods of 15 days | Total = 60 | Total = 59 | Total = 40 | |
| Wheat a - (3a) | 5th May to 20th May | 10 | 10 | 10 |
| 23rd June to 8th July | 10 | 10 | 8 | |
| 9th July to 24th July | 10 | 10 | 8 | |
| 9th Aug to 24th Aug | 10 | 8 | 7 | |
| 23rd Sept to 8th Oct | 10 | 9 | 7 | |
| 8th Oct to 23rd Oct | 10 | 0 | 0 | |
| Total = 6 periods of 15 days | Total = 60 | Total = 47 | Total = 40 | |
| Wheat b - (3bi) | 5th May to 20th May | 10 | 10 | 10 |
| 23rd June to 8th July | 10 | 8 | 8 | |
| 9th July to 24th July | 10 | 10 | 8 | |
| 9th Aug to 24th Aug | 10 | 7 | 7 | |
| 23rd Sept to 8th Oct | 10 | 10 | 7 | |
| 8th Oct to 23rd Oct | 10 | 7 | 0 | |
| Total = 6 periods of 15 days | Total = 60 | Total = 52 | Total = 40 |
From the 391 successful traps, 320 (40 traps from each field) were chosen for use in the data analysis (Table
The traps themselves were made out of 250 ml plastic cups. These had a depth of 10 cm and a rim diameter of 7.3 cm. They were part filled with ethylene glycol, and covered with wooden lids to prevent flooding and the capture of larger, non-target species. This left a gap of 2 cm between the trap rims and their lids.
The Carabidae were identified to species level using
Data Analysis
For each field, the data from 40 traps were combined, then the number of carabid species found in each field were recorded, along with their relative abundance (n). The annual activity density (ADa) (
ADa = ntot / US
Where:
ntot = the number of individuals sampled during the season
US = sum of us
us = trap * (gg/10)
trap = number of traps in each field
gg = the number of days the traps were set for.
The diversity of carabid species was calculated for each field using the Simpson's Diversity Index (D), which is presented here as the complement (1-D).
Carabid abundance and species richness were also calculated for each trap used in the data analysis. The resulting 40-trap data sets were tested using the Anderson-Darling test. This showed that the data sets were rarely normally distributed, even after transformation, meaning that nonparametric statistics were used for significance testing. Mann-Whitney U tests were used to determine the significance of differences between heterogeneous and homogeneous areas.
To compare between the different crop types, carabid abundance and species richness were again calculated for each trap. Then the data from both fields of each crop type were combined, resulting in four, 80-trap data sets, one for each crop type. Kruskal-Wallis tests were then used to determined the significance of differences between the cotton, maize, olive and wheat cultivations.
Results
Suppl. material
The total abundance (N), total annual activity density (ADa), species richness and diversity (1-D) of Carabidae in each field.
| Sampled Field - (Study Area) | Total Abundance (N) | Total Annual Activity Density (ADa) | Species Richness | Diversity (1-D) |
|---|---|---|---|---|
| Cotton a - (3a) | 9 | 0.150 | 5 |
0.86 |
| Cotton b - (3bii) | 71 | 1.183 | 6 | 0.60 |
| Maize a - (1a) | 892 | 14.869 | 11 | 0.48 |
| Maize b - (1b) | 897 | 14.950 | 7 | 0.34 |
| Olives a - (2a) | 105 | 1.750 | 11 | 0.69 |
| Olives b - (2b) | 47 | 0.683 | 14 | 0.97 |
| Wheat a - (3a) | 681 | 11.350 | 9 | 0.09 |
| Wheat b - (3bi) | 208 | 3.467 | 14 | 0.97 |
Checklist
Karya, Loutra Ipati, Mexiates
Sampling took place in cotton, maize, olive, and wheat fields. It was conducted by Anna Chapman (National and Kapodistrian University of Athens) and took place between the 5th of May and the 23rd of October 2007. All samples were preserved in alcohol and are now kept in the author's private collection.
Acinopus (Acinopus) picipes
Western Europe to Near East and Iran (
It digs burrows under stones and is mostly phytophagous (
Amara (Amara) aenea
From Macaronesia across Europe and the Mediterranean Region to Western Siberia (http://www.faunaeur.org/full_results.php?id=382134).
Xerophilous species, mainly inhabiting grassland, gardens, dunes and wasteland (
Amara (Amara) similata
Near transpalaearctic (
It prefers damp areas, riverbanks and water meadows (
Brachinus (Brachynidius) explodens
Prefers dry grassland and agricultural land, where it may be found in arable cultivations and alfalfa. It is one of the most common species of Carabidae to be found in cultivated areas in Eastern Europe (
Calathus (Bedelinus) circumseptus
Mediterranean Europe and parts of North Africa (
This species was rare in this study and was only found in the wheat field in the homogeneous area (n = 1).
Calathus (Calathus) korax
Endemic to Greece, but widespread within the country (
This species was found in the cotton field in the heterogeneous area (n = 1), the maize field in the heterogeneous area (n = 51), the olive grove in the homogeneous area (n = 4), the wheat field in the heterogeneous area (n = 1) and the wheat field in the homogeneous area (n = 5).
Calathus (Neocalathus) melanocephalus
Throughout Europe, to Western Asia and North Africa (
In agricultural areas, it is often found on arable land, pastureland and alfalfa. It is usually absent in fields with abundant weed cover (
Carabus (Oreocarabus) preslii
Greece, Italy and the Balkans (http://www.faunaeur.org/full_results.php?id=386874).
In this study, this species was only found rarely (n = 3) in the olive grove in the homogeneous area.
Carabus (Pachystus) graecus
Greece, Turkey, the Balkans and the Middle East (http://www.faunaeur.org/full_results.php?id=386941). Within Greece it is found on the mainland and Peloponnisos (
In this study, it was found in the olive grove in the heterogeneous area (n = 3), the olive grove in the homogeneous area (n = 7), the wheat field in the heterogeneous area (n = 3) and the wheat field in the homogeneous area (n = 12).
Carterus (Carterus) rufipes
Eastern European, the Mediterranean region, the Balkan Peninsula, the Caucasus, Asia Minor and the Near East (
It is a phytophagous and xerophilous species (
Carterus (Carterus) rotundicollis
The Western Mediterranean and the Balkan Peninsula (
It prefers open countryside (
Cylindera germanica
Europe and large parts of Asia (
Found on loamy soil, often on flood plains (
Dixus obscurus
The Balkans, Cyprus, Asia Minor, Iran, Iraq, the Caucasus and Southern Russia (
It was found in the olive grove in the heterogeneous area (n = 14) and the wheat field in the heterogeneous area (n = 3).
Harpalus (Harpalus) atratus
Europe (except the north) and the Balkans, where it prefers foothills to alpine regions (
It is a mesoxerophilous and polyphagous species, which prefers to live in forested areas (
Harpalus (Harpalus) dimidiatus
Western Europe to the Caucasus and the Middle East (
A species of dry grassland, which prefers moderate temperatures and humidity levels (
Harpalus (Harpalus) smaragdinus
Most of Europe (except the north), Asia Minor, east to Western Siberia and Western China, where it prefers plains to mountains (
It found rarely in the olive grove in the homogeneous area (n = 1) and the wheat field in the heterogeneous area (n = 1).
Harpalus (Pseudoophonus) rufipes
From the Azores, across Europe, to North Africa and Western China (
It is polyphagous and prefers open, dry habitats and light soils. It is most often found on arable land (
Microlestes luctuosus
Southern Europe and Southwest Asia, widespread and common in Greece (
It prefers warm, dry places (
Olisthopus fuscatus
Southern and Western Europe, as well as the Near East (http://www.faunaeur.org/full_results.php?id=379434).
Zoophagous (
Ophonus (Hesperophonus) azureus
Northwestern Africa, Northern, Central and Southern Europe, the Balkans, the Caucasus, Asia Minor and Northwestern China (
Consumes the seeds of common agricultural weed species such as Capsella bursa-pastoris, Taraxacum officinale and Cirsium arvense (
Ophonus (Ophonus) diffinis
From the Iberian Peninsular, through Southern and Central Europe, the Balkans, to the Near East and the Caucasus (
It is polyphagous, taking insect prey, but is also known to feed on the fallen seeds of plants in the Apiaceae family (
Pachycarus (Mystropterus) cyaneus
Greece, FYROM (Former Yugoslav Republic of Macedonia) Bulgaria and Turkey (
It lives in burrows underneath stones. A xerophilous species, preferring areas with sparse vegetation (
Pangus scaritides
Southern Russia, the Caucasus, Iran, Asia Minor, the Balkans as well as Southern and Central Europe (
This species was rare (n = 1) and was only found in the olive grove in the homogeneous area.
Poecilus cupreus
Europe, Asia Minor, Central Asia and Siberia (
It may be found in woodland, arable land, meadows, pastures and alfalfa fields. It is one of the most common carabid species of agricultural land in Central Europe. It feeds on species of Arachnida, Acari, Staphylinidae, Thysanoptera, Aphidoidea, other Hemiptera species, Cantharidae, Coccinellidae, Chrysopa and Lepidoptera larvae (
Pterostichus (Platysma) niger
Europe, Turkey, Iran, the Caucasus, Central Asia, Mongolia, Siberia and the Far East (
It is found in woodland, heathland and damp grassland (
Tapinopterus (Tapinopterus) taborskyi
Endemic to Greece and only ever found on Oiti mountain (
This species was found in the cotton field in the heterogeneous area (n = 2) and in the cotton field in the homogeneous area (n = 41). Both these areas were located close to Oiti.
Trechus (Trechus) quadristriatus
Europe, the Nearctic, the Near East and North Africa (http://www.faunaeur.org/full_results.php?id=384300). It is widespread and common in Greece (
This species was found in the maize field in the heterogeneous area (n = 1), the olive grove in the heterogeneous area (n = 1) and in the wheat field in the homogeneous area (n = 1).
Zabrus (Pelor) graecus
Greece, Bulgaria, FYROM and the Near East. Often found in Attica and on the near islands (
This species was found in the cotton field in the heterogeneous area (n = 3), the maize field in the heterogeneous area (n = 1), the maize field in the homogeneous area (n = 1), the olive grove in the heterogeneous area (n = 7), the olive grove in the homogeneous area (n = 1), the wheat field in the heterogeneous area (n = 17) and the wheat field in the heterogeneous area (n = 6).
Discussion
Neither the heterogeneous nor the homogeneous areas had consistently higher abundance, activity density, species richness, or diversity levels. This suggests that the level of heterogeneity of the study areas did not have a great influence on the carabid communities of the sampled fields. Areas with small field sizes, large amounts of non-cropped habitat and high land use diversity did not appear to benefit the Carabidae.
Additionally, there did not seem to be an association between any of the landscape metrics in Suppl. material
The results of this study do not agree with those reviewed by
A related issue is that too few land use types may have been sampled in this study. Heterogeneity is believed to enhance biodiversity, through different species being associated with different land use types, at different times in their lives (
Additionally, the need to compare closely situated areas, matched regarding other factors apart from their heterogeneity, meant that it was difficult to choose areas that differed greatly in all aspects of their heterogeneity. For example, although "land use diversity" was high and "land use similarity" was low in all of the heterogeneous areas, "land use richness" did not always follow this pattern. For the wheat comparison, "land use richness" was slightly higher in the homogeneous area, while for the cotton comparison "land use richness" was the same in both areas (Suppl. material
It is also possible, as mentioned by
Despite these issues, significant differences were seen in some of the comparisons between heterogeneous and homogeneous areas (Following Subsections). These results are interesting as they provide information about how the Carabidae within individual fields are affected by heterogeneity at the landscape level.
Relative Abundance
For the cotton fields, carabid abundance per trap was significantly higher in the homogeneous area (U = 1178, p = 0.0003). For the maize fields though, there was not a significant difference between the heterogeneous and homogeneous area (U = 831.5, p = 0.7642). The olive groves had significantly higher carabid abundance in the heterogeneous area (U = 586, p = 0.0404). The result of this comparison was one of the few that showed a positive influence of heterogeneity. For the wheat fields though, there was not a significant difference between the heterogeneous and homogeneous area (U = 693, p = 0.3077). Finally, when the data from all of the fields in each type of area were combined, there was no significant difference between heterogeneous and homogeneous areas (U = 12721 p = 0.9283).
On the whole, crop type appeared to have a greater influence on the Carabidae than did heterogeneity. The results of the Kruskal-Wallis test show that there was a highly significant difference in carabid abundance between the different crop types (H = 34, p = <0.0001). This was probably due to the specific microclimates created by each crop type and to the differences in husbandry that each crop required (
When the abundances of individual species were considered, significant differences were seen between some heterogeneous and homogeneous areas, but these were not consistent for all of the crop type comparisons. Relative abundance patterns also varied depending on the carabid species, with some species showing higher abundances in a heterogeneous area, and others in a homogeneous area. This may have been due to differences in dispersal ability, causing variation in they way individual species experienced heterogeneity (
For the cotton comparison, Pterostichus (Platysma) niger was found in significantly higher numbers in the homogeneous area (U = 1041, p = 0.0209). Then when the influence of crop type was examined, significantly greater numbers of this species were seen in the maize fields (H = 136.13, p = <0.0001). This may have been largely due to the maize fields being irrigated regularly throughout the summer, which would have provided favourable conditions for this species.
For the olive comparison, Microlestes luctuosus was found in significantly greater numbers in the heterogeneous area (U = 592, p = 0.0455). This species is known to prefer areas of tall vegetation (
For the maize comparison, Poecilus cupreus was found in significantly greater numbers in the heterogeneous area (U = 521, p = 0.0074). This may have been due to a preference for areas of non-cropped habitat at certain times of life (
For the cotton comparison, Tapinopterus (Tapinopterus) taborskyi was found in significantly greater numbers in the homogeneous area (U = 1050, p = 0.0164). This may suggest a preference for low heterogeneity levels. However, as this species has only ever been found on Oiti mountain (
For the maize comparison, Cylindera germanica was found in significantly higher numbers in the homogeneous area (U = 1199, p = 0.0001). This was probably because C. germanica is accustomed to living in agricultural areas (
The most common species in this study, Harpalus (Pseudoophonus) rufipes, did not show any significant differences between heterogeneous and homogeneous areas. This is probably due to it being a very common inhabitant of agricultural land (
Species Richness
For the cotton fields, there were significantly higher numbers of carabid species per trap in the homogeneous area (U = 1190, p = 0.0002), something that does not indicate a positive influence of landscape heterogeneity. For the maize and wheat fields, no significant differences were seen between heterogeneous and homogeneous areas (maize U = 673.5, p = 0.2263, wheat U = 759.5, p = 0.7039). For the olive groves too, there was not a significant difference between the heterogeneous and homogeneous area (U = 644.5, p = 0.1362). Additionally, when the data from all of the fields in each type of area were combined, there was no significant difference between heterogeneous and homogeneous areas (U = 13255, p = 0.5823).
The results of the Kruskal-Wallis test; however, showed that there was a highly significant difference in species numbers per trap between the different crop types (H = 90, p = <0.0001). The olive groves and the wheat fields had the highest overall richness levels of the four crop types. This may have been because these fields were organically farmed (
Diversity
Carabid diversity levels were not consistently higher in either the heterogeneous or the homogeneous areas. Nor was there a clear association between carabid diversity levels and the levels of any of the landscape metrics in Suppl. material
Neither were diversity levels consistently higher in any one crop type. Although overall diversity levels were highest in the olives groves, the least disturbed of all of the different cultivations. The highest diversity levels for individual fields were seen in the wheat and olive cultivations in the homogeneous areas. However, the lowest diversity level was seen in the wheat field in the heterogeneous area.
Again these results suggest that the level of heterogeneity had little influence on the Carabidae in the sampled fields. Although, as previously discussed, these findings may have been due to limitations imposed by this study's design.
Acknowledgements
References
- The Ground Beetles of Northern Ireland. Atlases of the Northern Ireland Flora and Fauna.1.Ulster Museum,246pp.
- Ground Beetles (Carabidae) of Greece.Pensoft,Sofia,394pp.
- Farmland Biodiversity: is Habitat Heterogeneity the Key?Trends in Ecology and Evolution18:182‑188. https://doi.org/10.1016/S0169-5347(03)00011-9
- I Coleotteri Carabidi per la valutazione ambientale e la conservazione della biodiversità. APAT, Manuali e linee guida.34.I.G.E.R. srl,Roma,240pp. [InItalian].
- What is Going on Between Aposematic Carabid Beetles? The Case of Anchomenus dorsalis (Pontoppidan 1763) and Brachinus sclopeta (Fabricius, 1792) (Coleoptera Carabidae).Ethology Ecology & Evolution18:335‑348. https://doi.org/10.1080/08927014.2006.9522700
- Fenología de las Espacies de Carábidos (Col. Carabidae) Más Abundantes en la Cuenca de Bembezar (NW. de la Provincia de Cordoba).Mediterranea Serie de Estudios Biológicos8:147‑163.
- Ground beetle abundance and community composition in conventional and organic tomato systems of California's Central Valley.Applied Soil Ecology11:199‑206. https://doi.org/10.1016/s0929-1393(98)00138-3
- Loss of Habitats and Changes in the Composition of the Ground and Tiger Beetle Fauna in Four West European Countries since 1950 [Coleoptera: Carabidae, Cicindelidae].Biological Conservation48:277‑294. https://doi.org/10.1016/0006-3207(89)90103-1
- Ecological Processes That Affect Populations in Complex Landscapes.Oikos65(1):169. https://doi.org/10.2307/3544901
- Agricultural Statistics. Main Results -2010-11. Eurostat Pocket Books.Office for Official Publications of the European Communities.,Luxembourg.,228pp. https://doi.org/10.2785/3341
- The Role of Research and Development in the Evolution of a ‘Smart’ Agri-environment Scheme.Aspects of Applied Biology67:253‑264.
- Investigating the effects of crop type, fertility management and crop protection on the activity of beneficial invertebrates in an extensive farm management comparison trial.Annals of Applied Biology155(2):267‑276. https://doi.org/10.1111/j.1744-7348.2009.00337.x
- Consequences of the Cessation of 3000 Years of Grazing on Dry Mediterranean Grassland Ground-Active Beetle Assemblages.Comptes Rendus Biologies331:532‑546. https://doi.org/10.1016/j.crvi.2008.04.006
- Functional Landscape Heterogeneity and Animal Biodiversity in Agricultural Landscapes.Ecology Letters14:101‑112. https://doi.org/10.1111/j.1461-0248.2010.01559.x
- The Effects of Agricultural Practices on Carabidae in Temperate Agroecosystems.Integrated Pest Management Reviews5(2):109‑129. https://doi.org/10.1023/A:1009619309424
- Migrants in Rural Greece.Sociologia Ruralis43:167‑184. https://doi.org/10.1111/1467-9523.00237
- Carabid Beetles in Sustainable Agriculture: a Review on Pest Control Efficacy, Cultivation Impacts and Enhancement.Agriculture, Ecosystems and Environment74:187‑228. https://doi.org/10.1016/S0167-8809(99)00037-7
- Farmers and Agricultural Policy in Greece since the Accession to the European Union.Sociologia Ruralis37:270‑286. https://doi.org/10.1111/j.1467-9523.1997.tb00050.x
- The Carabidae (Ground Beetles) of Britain and Ireland. Handbooks for the Identification of British Insects. Vol. 4 Part 2 (2nd Ed.).Royal Entomological Society and the Field Studies Council,252pp.
- Relationships of Natural Enemies and Non-Prey Foods.Progress in Biological Control7:143‑165.
- FRAGSTATS: Spatial Pattern Analysis Program for Categorical Maps. Computer Software Program.University of Massachusetts.
- Zur Biologie und Ökologie der ripicolen Carabiden Bembidion femoratum Sturm und B. punctulatum Drap.II. Die Substratbindung. Zoologische Jahrbuecher Systematik111:369‑383.
- Landscape Complexity and Farming Practice Influence the Condition of Polyphagous Carabid Beetles.Ecological Applications11:480‑488. https://doi.org/10.1890/1051-0761(2001)011[0480:LCAFPI]2.0.CO;2
- Arthropods in the Mounds of Mole Rats, Spalax ehrenbergi Superspecies, in Israel.Ecologia Mediterranea31:5‑13.
- Weeds in Agricultural Landscapes. A Review.Agronomy for Sustainable Development31:309‑317. https://doi.org/10.1051/agro/2010020
- Effects of Bio-dynamic, Organic and Conventional Farming on Ground Beetles (Col. Carabidae) and Other Epigaeic Arthropods in Winter Wheat.Biological Agriculture & Horticulture12(4):353‑364. https://doi.org/10.1080/01448765.1996.9754758
- Hedges III. The Effect of Removal of the Bottom Flora of a Hawthorn Hedge on the Carabidae of the Hedge Bottom.Journal of Applied Ecology5:125‑139. https://doi.org/10.2307/2401278
- Occurrence and Fauna Composition of Ground Beetles in Wheat Fields.Journal of Central European Agriculture11:423‑432. https://doi.org/10.5513/JCEA01/11.4.855
- Pachycarus (Mysteropterus) cyaneus (Dejean 1825) - Species of Ground Beetles (Coleoptera, Carabidae) New for Bulgaria‘s Fauna.Polish Journal of Entomology77:63‑66.
- The Distribution and Abundance of Predatory Coleoptera Overwintering in Field Boundaries.Annals of Applied Biology106:17‑21. https://doi.org/10.1111/j.1744-7348.1985.tb03089.x
- Effects of Agricultural Diversification on the Abundance, Distribution, and Pest Control Potential of Spiders: a Review.Entomologia Experimentalis et Applicata95:1‑13. https://doi.org/10.1046/j.1570-7458.2000.00635.x
- Carabid Beetles in Their Environments.Springer,Berlin/Heidelberg,372pp. https://doi.org/10.1007/978-3-642-81154-8
- Nota Sobre el Género Calathus Bonelli, 1810, en la Península Ibérica (Coleoptera, Carabidae).Bulletin de la Société Entomologique de France111:51‑57.
- Tiger Beetles Ground Beetles. Illustrated Key to the Cicindelidae and Carabidae of Europe.Josef Margraf,Aichtal,488pp.
- Landscape Perspectives on Agricultural Intensification and Biodiversity – Ecosystem Service Management.Ecology Letters8:857‑874. https://doi.org/10.1111/j.1461-0248.2005.00782.x
- Trophic Level Modulates Carabid Beetle Responses to Habitat and Landscape Structure: a Pan-European Study.Ecological Entomology35:226‑235. https://doi.org/10.1111/j.1365-2311.2010.01175.x
- Diversity and the Main Ecological Requirements of the Epigeic Species of Carabidae (Coleoptera, Carabidae) in the Sun Flower Ecosystem, Broscauti (Botosani County).Analele Stiintifice ale Universitatii „AI. I. Cuza” Iasi, s. Biologie AnimalaLIV:1‑9.
- Diversity of Butterflies in the Agricultural Landscape: the Role of Farming System and Landscape Heterogeneity.Ecography23:743‑750. https://doi.org/10.1111/j.1600-0587.2000.tb00317.x