|
Biodiversity Data Journal :
Taxonomic Paper
|
|
Corresponding author: Andrew D. F. Simon (adfsimon@imerss.org)
Academic editor: Yasen Mutafchiev
Received: 02 Oct 2021 | Accepted: 21 Jan 2022 | Published: 10 Mar 2022
This is an open access article distributed under the terms of the CC0 Public Domain Dedication.
Citation:
Simon ADF, Adamczyk EM, Basman A, Chu JWF, Gartner HN, Fletcher K, Gibbs CJ, Gibbs DM, Gilmore SR, Harbo RM, Harris LH, Humphrey E, Lamb A, Lambert P, McDaniel N, Scott J, Starzomski BM (2022) Toward an atlas of Salish Sea biodiversity: the flora and fauna of Galiano Island, British Columbia, Canada. Part I. Marine zoology. Biodiversity Data Journal 10: e76050. https://doi.org/10.3897/BDJ.10.e76050
|
|
Abstract
Background
Based on records dating from 1859 to 2021, we provide an overview of the marine animal diversity reported for Galiano Island, British Columbia, Canada. More than 650 taxa are represented by 20,000 species occurrence records in this curated dataset, which includes dive records documented through the Pacific Marine Life Surveys, museum voucher specimens, ecological data and crowd-sourced observations from the BC Cetacean Sightings Network and iNaturalist.
New information
We describe Galiano Island's marine animal diversity in relation to the Salish Sea's overall biodiversity and quantify the proportional contributions of different types of sampling effort to our current local knowledge. Overviews are provided for each taxonomic group in a format intended to be accessible to amateur naturalists interested in furthering research into the region's marine biodiversity. In summary, we find that the Pacific Marine Life Surveys, a regional community science diving initiative, account for 60% of novel records reported for Galiano Island. Voucher specimens account for 19% and crowd-sourced biodiversity data 18% of novel records, respectively, with the remaining 3% of reports coming from other sources. These findings shed light on the complementarity of different types of sampling effort and demonstrate the potential for community science to contribute to the global biodiversity research community. We present a biodiversity informatics framework that is designed to enable these practices by supporting collaboration among researchers and communities in the collection, curation and dissemination of biodiversity data.
Keywords
Salish Sea, Galiano Island, marine zoology, biodiversity, biodiversity informatics, inclusive design, open data, community science, community-integrated research
Introduction
Galiano Island is located on the northwest coast of North America (
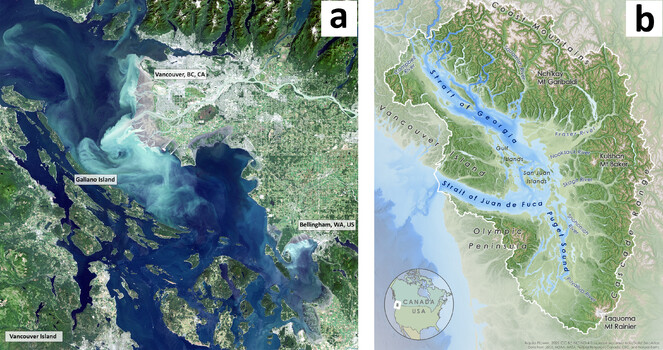
Study area: Galiano Island, British Columbia, Canada (Salish Sea bioregion)
a. Satellite image showing Galiano Island in relation to major population centres. The Salish Sea, as a dynamic estuarine ecosystem, is illustrated in this enhanced image highlighting sediments from the Fraser and Nooksack Rivers. Imagery from Landsat-7 EMT+, 30 July 2000. Projection: UTM Zone 10 NAD83. Scale 1:200,000. – Image by Galiano Conservancy Association.
b. Physical geography reference map for the Salish Sea bioregion. The Salish Sea bioregion includes both marine waters and their upland watersheds. These boundaries technically distinguish the Salish Sea from the Puget Sound/Georgia Basin marine ecoregion (sensu
Galiano Island is part of the San Juan Archipelago, which spans the Canada-USA border in the rain shadow of the Olympic Mountains and Vancouver Island Ranges. This sheltered subregion of the Salish Sea has a Mediterranean-type climate that supports some of Canada’s most imperilled ecosystems, including the highest density of threatened species in British Columbia (
Given the increasing threats that anthropogenic climate change and development present for the Salish Sea (
This article is the first instalment in a five-part series documenting Galiano Island’s flora and fauna. Each part in this series is intended to: a) establish a formal biodiversity baseline for Galiano Island; and b) elaborate an open source framework for organising, evaluating, analysing and sharing diverse sources of biodiversity data, which may be adapted as the basis for an atlas of Salish Sea biodiversity. Thus, while this project is being advanced with a focus on Galiano Island, it is intended to serve a broader purpose in the field of biodiversity informatics: to create a more inclusive and systematic framework for biodiversity research and monitoring. Community science has become an increasingly important source of data for international biodiversity monitoring efforts (
Study Area Description
The Salish Sea is a complex and dynamic estuarine ecosystem, within which Galiano Island is centrally situated (Fig.






Marine habitat diversity around Galiano Island.
b: Marine habitat diversity at Sumnuw (Montague Harbour) includes shell beaches, tidal mudflats and salt marsh, the product of thousands of years of Indigenous landscape engineering
c: Dragon rock: a sandstone reef off the southwest coast of Parker Island, near Galiano Island
d: The cloud sponge (Aphrocallistes vastus) forms an extensive deep-sea reef off the southeast coast of Galiano Island, providing habitat for animals, such as the brown box crab (Lopholithodes foraminatus)
e: Juvenile copper rockfish (Sebastes caurinus) take refuge in an eelgrass meadow laden with diatoms, bacteria and other micro-organisms
f: A bed of young bull kelp (Nereocystis luetkeana) interspersed with the introduced Japanese wireweed (Sargassum muticum) on a rocky shoreline nearby Collinson Point, Galiano Island.
Galiano Island's coastal environment has a long history of habitation and cultural modification by Coast Salish Indigenous peoples, who intentionally enhanced the productivity of these rich marine ecosystems (
Materials and methods
Data curation
This dataset was curated through a systematic evaluation of taxonomic summaries and numerous catalogues of species occurrence data. We began by compiling unique taxa from source catalogues and resolving their taxonomy with reference to the World Register of Marine Species (WoRMS) (
The data sources contributing to this baseline dataset have been normalised, aligned, corrected, synthesised and rendered into visualisations by a set of open source data processing scripts written in JavaScript. The algorithms developed to facilitate these workflows conserve memory of all modifications to the dataset, including changes to taxonomy, georeferencing and typographic errors. Georeferencing was reviewed and corrected with reference to the best available metadata using QGIS version 3.1.0 (
Data processing algorithms
The curation of this dataset was facilitated by algorithms that operate in the following steps:
- The columns of each source catalogue, imported as CSV, are mapped with a common core of fields drawn from a subset of the Darwin Core standard (
Wieczorek et al. 2012 ). - The taxon name is mapped with a core taxonomic backbone by means of a taxon resolution file which resolves preferred taxon names and accounts for typographical errors.
- A dataset id is assigned to every source catalogue, which are then combined into a single master catalogue.
- This catalogue is filtered to include only the taxa of interest—marine fauna.
- Private or obscured coordinates held in project-specific fields are copied into the principal georeferencing fields.
- A patch of georeferencing corrections is applied to the resulting coordinates, together with curatorial notes justifying the corrections.
- The resulting observations are filtered by the polygon representing the project area.
- The resulting output produces two consolidated CSV files: a catalogue of all observations and a master summary file.
- The master summary file is then divided into phyla for curation by subject matter experts—these are exported into a Google Sheets representation where experts may edit them live.
- Subject matter experts add curation notes and resolve taxonomic discrepancies, the results of which are checked against authorities listed in WoRMS (
WoRMS Editorial Board 2021 ). - After curation, the Google Sheets are converted back into CSV, mirrored into the GitHub repository, re-integrated and compared with the original summary produced at Step 8.
- Where appropriate, discrepancies between the summaries produced at Step 8 and Step 11 are circulated among the managers of the source catalogues to incorporate corrections. Residual discrepancies are then fed into the taxon resolution file at Step 2.
- The process is re-run from Step 2 until repeated passes give rise to no further discrepancies at Step 12.
The files output at Step 8 of the pipeline form the basis of the map-based data visualisations referenced from this paper, as well as our Darwin Core data submission to the Global Biodiversity Information Facility (
Rationale
Our approach enhances common data cleaning practices by making minimal modifications to the source catalogues as they are incorporated into the dataset. Instead, source catalogues are retained in their original form and any transformations required by the data cleaning and normalisation process are encoded as a set of reversible patches or lenses applied to the data. In addition to supplying thorough documentation of our data-cleaning workflow, in line with best practices (
For example:
- Taxonomic adjustments and spelling corrections pertaining to records in catalogues which we are unable to amend (e.g. Canadian Museum of Nature collections) are held in a taxon swaps file which is re-applied to the catalogues whenever they are updated. This process results in an institutional memory of the relationships between preferred taxonomies and those of contributors, while at once creating a log of commonly encountered typographical errors. Memory of such changes can be applied in the resolution of future datasets, helping to reduce redundancy in curatorial processes.
- Georeferencing corrections to historical records of poor spatial resolution are held in a georeferencing patch file, which may be re-applied to source catalogues when they are updated, without requiring the original source to accept our corrections.
- The iterative process described in Step 13 accommodates updates from ongoing observations and corrected upstream sources, as well as taxon-directed updates from subject matter experts, while ensuring that these updates do not introduce new errors into the system.
Data resources
Scientific records of Galiano Island’s marine fauna date back over a century to specimens collected by Alexander Agassiz in 1859 (
The Pacific Marine Life Surveys represent the greatest proportion of data included in this study, with 16,150 species occurrence records accounting for 504 taxa (species, subspecies and species complexes), 241 of which are unique reports (Fig.
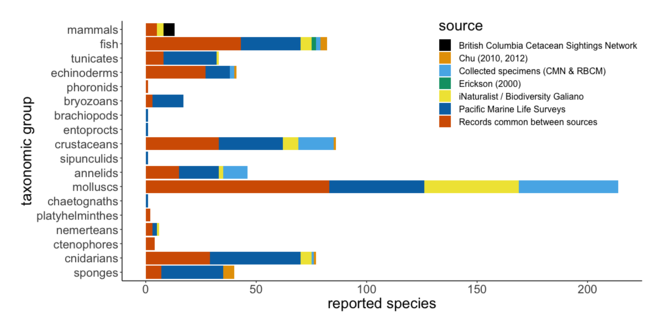
Bar plot summarising unique species reports by data source. Taxonomic overlap between catalogues includes 263 species reports that are common between sources. Note: voucher specimens representing two cnidarian species, cited by
Table
Summary of marine animal diversity reported for Galiano Island vis-à-vis global and regional biodiversity estimates. Regional diversity is estimated for the Pacific coast of North America (Pacific NA), the Northeast Pacific (NEP), coastal British Columbia (BC) and the Salish Sea, depending on the available data. See the taxonomic summaries below for sources of diversity estimates.
|
Group |
Global diversity |
Regional diversity |
Local diversity (Galiano Island) |
Note |
|
Sponges (Porifera) |
9,452 species |
300–400 species (BC); 70 species (Salish Sea) |
40 species |
|
|
Cnidarians (Cnidaria) |
12,000 species |
600 species (Pacific NA); 200 species (BC) |
77 species |
|
|
Ctenophores (Ctenophora) |
150–200 species |
32 species (Pacific NA); 13 species (Salish Sea)* |
4 species |
*only 5 or 6 taxa commonly occur in the Salish Sea |
|
Ribbon worms (Nemertea) |
> 1,300 species |
41 species (BC); 30 species (Salish Sea) |
6 species |
|
|
Flatworms (Platyhelminthes) |
13,000 marine species* |
> 170 species (BC) |
2 species |
*this estimate does not account for cryptic parasitic species (e.g. Trematodes, (> 18,000 taxa) |
|
Arrow worms (Chaetognatha) |
130 species |
6 species (NEP); 4 species (BC) 1–2 species (Salish Sea) |
?* |
*chaetognaths in the Galiano Island record remain undetermined to species |
|
Molluscs (Mollusca) |
49,000 species* |
780 species (BC) |
214 species |
*global estimate incl. both terrestrial and marine spp; regional and local richness incl. only marine spp. |
|
Ringed worms (Annelida) |
> 13,000 species |
> 450 species (BC); 860 species (Salish Sea)* |
46 species |
*known regional diversity vastly underestimates true richness |
|
Peanut worms (Sipuncula) |
150 species |
8 species (NEP) |
1 species |
|
|
Crustaceans (Crustacea) |
52,000 species* |
900 species (BC) |
86 species |
*global estimate incl. both terrestrial and marine spp; regional and local richness incl. only marine spp. |
|
Nodding-heads (Entoprocta) |
253 species |
11 species (BC) |
1 species |
|
|
Lampshells (Brachiopoda) |
400 species |
7 species (BC); 3 species (Salish Sea) |
1 species |
|
|
Bryozoans (Bryozoa) |
> 6,000 species |
210–260 species (BC) |
17 species |
|
|
Horseshoe worms (Phoronida) |
13 species |
6 species (NEP) |
1 species |
|
|
Echinoderms (Echinodermata) |
> 7,000 species |
111 species (BC) |
41 species |
|
|
Tunicates (Tunicata) |
2,815 species |
71 species (BC) |
33 species |
|
|
Ray-finned and cartilaginous fish (Actinopterygii & Chondrichthyes)* |
> 33,000 species > 1,100 species |
> 1,500 species (Pacific NA); 400 species (BC) |
82 species |
*diversity estimates for Actinopterygii and Chondrichthyes are combined in regional and local summaries |
|
Mammals (Mammalia) |
6,400 species |
22 species (Salish Sea) |
13 species |
Taxonomic summaries
Phylum
[3 classes: 15 orders: 27 genera: 40 species]
The name ‘Porifera’ derives from Modern Latin and means, literally, “bearing pores”, referring to the pores that perforate the body wall of these organisms.
Porifera are commonly known as sponges and have the simplest body plans of all multicellular animals, with no tissues or organs. Currently there are 9,452 valid species recognised worldwide, including representatives from both fresh and saltwater habitats (
Most sponges are filter feeders that tend to thrive in strong current or wave-swept habitats. In tidal passages along the coast of British Columbia, sponges are often the most conspicuous and abundant sessile invertebrates present (Neil McDaniel, pers. obs.). Given that it is difficult to census the biodiversity found in areas with strong currents, additional species have likely gone undocumented from channels around Galiano Island. The homosceleromoph sponge Oscarella sp., for example, is known to occur in inlets of Vancouver Island and would likely be found in deeper waters around Galiano Island with further search effort. This dataset also includes three undescribed sponge species, which is likely a small percentage of the undescribed species in this region. Indeed, new sponge species are frequently described based on specimens sampled in deeper waters (
The glass sponges Aphrocallistes vastus Schulze, 1886 (cloud sponge) (Fig.

Uncertainty regarding sponge diversity primarily results from their cryptic nature, which demands specialised knowledge of their skeletal structures and advanced microscopy techniques to confirm species. Most sponges are characterised by their calcareous or siliceous spicules. The abundance, morphology and arrangement of these spicules, along with the form, texture and colour of the sponge, are used to identify specimens with reference to previously-described species. Austin and Ott (
Phylum
[3 classes: 13 orders: 42 families: 64 genera: 77 species]
‘Cnidaria’ is the Latinised plural form of the Greek κνίδη (knidē), which means “nettle.”
Cnidaria comprise 12,000 extant invertebrate species which primarily occur in marine environments (
Cnidarians occur widely throughout the marine environment, with medusae present in pelagic and polyps in benthic habitats. While field identification is reliable in some cases, many taxa, especially hydrozoans, require microscopy for reliable determination. Owing to these limitations, certain groups, including anemones and corals, are well represented in the Galiano Island record, whereas others, such as hydroids and hydrocorals and the smaller scyphozoans, are not. The deeper-dwelling soft corals (gorgonians) are also under-represented due to sampling biases toward shallower waters.
In this dataset, we report the recently-described hydroids Leuckartiara longicalcar Schuchert, 2018 (Fig.

Other reported taxa require further study to resolve their taxonomy. For instance, recent molecular analysis of Aequorea in the Northeast Pacific show they belong to a complex (here reported as Aequorea victoria s. lat.), with many other taxa potentially included within this clade (Henry Choong and Wyatt Patry, pers. comm. 2020). Within Alcyonium, two taxa are recognised in the region, though they have yet to be circumscribed. Euphysa also likely represents a complex, the diversity of which has yet to be determined in our region (Claudia Mills, pers. comm. 2020).
The alternation between polypoid and medusoid generations has historically resulted in a great deal of confusion in the classification of cnidarians. Hydroids, in particular, have received little recent taxonomic attention, with the most substantive revisions dating to
Phylum
[2 classes: 3 orders: 4 families: 4 genera: 4 species]
‘Ctenophora’ means “comb-bearing”—deriving from the Greek κτείς (kteis), meaning "comb", and -φορος (-fóros), a suffix meaning "carrying”.
Ctenophora (comb jellies) are a phylum of predatory marine invertebrates that are characterised by having eight rows of ciliated plates for locomotion. The phylum is divided into two classes containing 150–200 extant species worldwide (

Most ctenophores are pelagic species, with those most well known occurring near the ocean surface and those less understood found at greater depths. Ctenophores possess sticky cells called colloblasts to capture prey and are are highly diverse in morphology (e.g. egg-shaped cydippids, flat and generally combless platyctenids and large-mouthed beroids). The phylogenetic position of Ctenophora in relation to other phyla is still debated and their taxonomy is in dire need of revision (
Phylum
[3 classes: 3 orders: 4 families: 4 genera: 6 species]
‘Nemertea’ is the Modern Latin (plural) form of the Greek νημερτής (Nēmertēs)—the name of a sea nymph.
Nemertea, also known as ribbon worms, are a phylum of soft-bodied and unsegmented invertebrate animals. Over 1,300 extant species, mostly found in marine environments, have been described globally (
Most nemerteans live in benthic environments, lurking in crevices beneath stones, shells and at the bases of algae or sessile animals. They are largely predatory or scavenging animals, feeding on annelids, clams and crustaceans, though some are commensalists that live within the mantle cavity of molluscs where they feed on micro-organisms filtered out by their hosts (
The key diagnostic features of Nemertea are proboscis characteristics: for example, whether it is armed or unarmed or split into regions. This makes identification of many species difficult as it requires examination of internal anatomy. Some species, such as Tubulanus sexlineatus (Coe, 1904) (six-lined ribbon worm) (Fig.

Phylum
[1 class: 1 order: 2 families: 2 genera: 2 species]
‘Platyhelminthes’ is rooted in the Greek πλατύ (platy), meaning “flat”, and ἑλμινθ- (helminth-), meaning “worm”.
Platyhelminthes (flatworms) are a phylum of dorsoventrally flattened, bilaterally symmetrical invertebrates, most of which are found in marine and freshwater environments (
Given the region's high diversity of cryptic free-living and parasitic flatworm species, expert search effort is necessary to gain a comprehensive understanding of this understudied phylum. The larger flatworm species in the order Polycladia are the most likely to be found diving or in intertidal areas. However, even with this more visible group, species identification can be difficult, requiring the fixing, cleaning and sectioning of specimens to examine the microscopic details of copulatory organs and other internal anatomy. Only the largest and most distinctly patterned taxa, such as Eurylepta leoparda Freeman, 1933 (spotted flatworm) (Fig.

Phylum
[1 class]
The name ‘Chaetognatha’ derives from the Greek χαίτη (khaítē), meaning “bristle”, and γνάθος (gnáthos), meaning “jaw”.
Chaetognatha (arrow worms) are a small phylum of predatory animals, all of which are marine, including 130 extant species in the class Sagittoidea (

Chaetognaths are active predators, grasping their prey with two sets of rigid hooks located at the sides of the head, hence the common name “bristle-mouth.” These predatory animals are chiefly pelagic, occurring in the open ocean. Appearing like a glass arrow in the water, chaetognaths swim by flexing in the middle of the body, which is supported by a hydrostatic skeleton. The lateral fins on either side of the body appear to aid buoyancy, but are not involved in movement. As few species are present locally, students may disregard more cryptic characteristics and distinguish species by focusing on the arrangement of lateral fins and eye colouration (
Phylum
[5 classes: 31 orders: 101 families: 151 genera: 214 species]
'Mollusca' is rooted in the Modern Latin 'mollusca', the neuter plural form of 'molluscus', from 'mollis', meaning “soft”.
With about 49,000 extant species (
Mollusc inventories have historically been limited by outdated taxonomy and other challenges, leaving much hidden diversity that remains to be discovered in the region. As with other groups, European species concepts have historically been misapplied to Northeast Pacific taxa among Mollusca. One example is the aeolid nudibranch Cuthona pustulata (Alder & Hancock, 1854), originally described from the Northeast Atlantic in 1854. Specimens collected under this name from Porlier Pass off Galiano Island by Sandra Millen in 1982 (RBCM 983-00026-001) have since been described as Zelentia nepunicea Korshunova, Fletcher, Lundin, Picton & Martynov, 2018 (pimpled aeolid, Fig.

While many nudibranch species revealed through genetic methods can be determined based on morphology, other cryptic taxa often cannot be reliably identified by morphology alone, requiring molecular analysis for confirmation. Locally occurring genera currently under taxonomic revision include Crepidula Lamarck, 1799 and Vermetus Daudin, 1800. Many more taxonomic issues remain to be resolved in future studies.
The cold, rich waters surrounding Galiano Island are home to many remarkable molluscs, including Enteroctopus dofleini (Wôlker, 1910) (giant Pacific octopus), the largest octopus in the world. The largest chiton in the world, Cryptochiton stelleri (von Middendorff, 1847) (gumboot chiton), is also known to the region and is reported for Galiano Island. Introduced bivalves present around Galiano Island include Magallana gigas (Thunberg, 1793) (Pacific oyster), Mya arenaria Linnaeus, 1758 (soft-shelled clam), Nuttalia obscurata (Reeve, 1857) (purple mahogany clam) and Ruditapes philippinarum (Adams & Reeve, 1850) (Japanese littleneck), all of which are now prolific in the Northeast Pacific. Introduced gastropods present include Batillaria attramentaria (G.B. Sowerby II, 1855) (Japanese false cerith) and Myosotella myosotis (Draparnaud, 1801) (mouse-eared snail).
Species that will likely be uncovered with further search effort include the threatened native Ostrea lurida (Carpenter, 1864) (Olympia oyster) and Penitella penita (Conrad, 1837) (flat-tip piddock). Numerous families of minuscule “micro-molluscs” are entirely unrepresented in the Galiano Island record. A diverse chiton fauna may also be under-represented locally, with 39 species in six families known from the intertidal zone to moderate diving depths (< 30 m) in the Salish Sea (
Molluscs were traditionally identified based on shell and other morphological characters. However, modern taxonomic treatments generally rely on genetics in addition to morphology, habitat and host species. Many snails also lay diagnostic egg cases. Regional references for Mollusca include:
Phylum
[2 classes: 6 orders: 15 families: 39 genera: 46 species]
‘Annelida’ was coined in Modern Latin by the French naturalist Jean-Baptiste Lamarck from the French 'annélide', deriving from 'annelés', meaning “ringed ones” (from the Latin 'anulus', for “little ring”).
Annelida are a large phylum of invertebrates, commonly referred to as ringed or segmented worms, comprising over 13,000 extant species (
Over 450 polychaete species are reported by
Annelid life history is exceedingly diverse, exhibiting virtually all feeding modes, including suspension feeding, deposit feeding, scavenging, herbivory, and carnivory. This diversity is mirrored in their pervasive distribution throughout most benthic and pelagic marine environments. Gaps in our knowledge of Galiano Island’s annelid diversity partly reflect the challenges that have historically limited our broader understanding of the region’s diversity. Taxonomists working in the region in the past largely relied on European references and only assigned new names for taxa with strikingly different features. However, the characters defining organisms within this challenging group are often very small and difficult to ascertain without resorting to dissection or microscopic examination. Furthermore, the vast majority of polychaetes lie hidden in bottom sediment or among sessile organisms and are rarely noticed by casual observers. Specialised equipment, such as benthic grabs, dredges, sediment air lifts, plankton tows and settlement plates, are needed to capture more than the few conspicuous species seen by beach-goers and divers.
The majority of taxa reported in this dataset are polychaetes, though a few clitellates are also represented. Many of the names reported reflect the ongoing challenges facing those studying this group. Uncertainty is indicated by older reports, such as Chaetopterus variopedatus (Renier, 1804) and Nereis zonata Malmgren, 1867 (here reported as the Chaetopterus variopedatus complex and Nereis cf zonata), which refer to European concepts in which undescribed native species likely remain hidden. Even conspicuous species, such as tubeworms in the genus Myxicola (Fig.

Polychaete morphology is highly variable, yet their body plan generally consists of an elongated and segmented body. It is the differentiation of their body into segments and the appendages (gills, parapodia, chaetae, cirri, palps etc.) attached to these segments that enables the morphological classification of families, genera and species. Regional references treating annelids include
Phylum
[1 class: 1 order: 1 family: 1 genus: 1 species]
‘Sipuncula’ is the Modern Latin plural form of the genus Sipunculus, based on the Latin 'siphunculus', meaning “small tube.”
Sipuncula (peanut worms) are a small phylum of bilaterally symmetrical, unsegmented marine worms, represented by 150 species worldwide (

Due to their burrowing habit, sipunculids are obscure organisms easily overlooked by divers and casual beachcombers. Some Sipuncula inhabit rock crevices or similar protected areas, while others are found in sand or mud. With additional search effort, other taxa likely to be uncovered in the waters around Galiano Island include Phascolosoma (Phascolosoma) agassizii Keferstein, 1866 and Themiste (Themiste) pyroides (Chamberlin, 1919). Identification relies on features of the tentacles which tend to be drawn into the body when contracted, hence the common name “peanut worms.” Regional references to the sipunculids can be found in
Subphylum
[3 classes: 7 orders: 32 families: 52 genera: 86 species]
The name ‘Crustacea’ is rooted in the Modern Latin 'crusta', meaning “shell.”
Crustacea are a highly diverse subphylum of Arthropoda that contains over 52,000 described terrestrial, marine and freshwater species (
Many crustaceans are difficult to identify without microscopy, contributing to biases in the local record. Locally, ichthyostracan crustaceans are vastly under-represented and cladocerans and ostracods remain unrepresented to date. As of yet, there are no records of hoplocarids (mantis shrimps), ascothoracids or tantulocarids occurring within British Columbia. The latter two groups will likely be discovered with further investigation; hoplocarids, however, have never been observed north of southern California and are unlikely to occur in the region. Leptostracans are not well described, though at least one species from the genus Nebalia is known to the Northeast Pacific (
Numerous non-native crustaceans are known to the region, though only one introduced species is currently represented in the Galiano Island record. The introduced Caprella mutica Schurin, 1935 (Japanese skeleton shrimp) (Fig.

References for regionally-occurring marine crustaceans include: amphipods (
Phylum
[1 class: 1 order: 1 family: 1 genus: 1 species]
‘Entoprocta’ derives from the Greek ἐντός (entos), meaning “inside”, and πρωκτός (prōktos), meaning “anus.”
Entoprocta (nodding-heads) are a small phylum of mainly sessile marine invertebrates that comprises 253 described species worldwide (

Entoprocts are filter feeders, trapping small particles in the mucus secreted by their tentacles. An understudied group, they are easily overlooked because of their diminutive size (< 1 mm) and the superficial similarities they share with the Cnidaria and Bryozoa (
The body plan of entoprocts consists of a cup-like calyx that bears a ring of ciliated tentacles called the lophophore, which is attached on its dorsal surface to the substrate by a long, thin stalk or pedicel. Entoprocta have their anus inside their ring of tentacles, while bryozoans (formally known as the Ectoprocta), have their anus outside of their ring of tentacles. Regionally occurring Entoprocta are treated by
Phylum
[1 class: 1 order: 1 family: 1 genus: 1 species]
‘Brachiopoda” is formed of the Ancient Greek βραχίων (brakhíōn), meaning “arm”, and πούς (poús), for “foot”.
Brachiopods (lampshells) are a phylum of shelled marine animals found in intertidal and subtidal waters, represented by over 400 species worldwide (

Most lampshells live at great depths and are thus seldom encountered. Laqueus californicus is known to occur in mass aggregations (945 m-2) at depths as deep as 700 m, which is likely why this species has eluded search efforts to date (
Traditionally, brachiopods were classified into two major groups, based on whether their valve hinge structures are articulate or inarticulate. Shells are orientated in a dorsal-ventral manner, closely resembling bivalve molluscs, the shells of which are lateral in orientation. Yet, whereas bivalves are symmetrical between valves (valves mirror each other), the plane of symmetry in brachiopods cuts through the middle of their valves (valves do not mirror each other). In contrast to bivalves, brachiopods are also typically attached to the substrate by a stalk. They share their feeding organ, the lophophore, in common with the Bryozoa, Entoprocta and Phoronida. Regional accounts of Brachiopoda include
Phylum
[2 classes: 3 orders: 13 families: 15 genera: 17 species]
‘Bryozoa’ derives from the Ancient Greek βρύον (brúon), “moss”, and ζῷα (zôia), which means “animals.”
Bryozoans, or ‘moss animals’, are colonial animals that are composed of many connected individuals, globally represented by over 6,000 species (
Bryozoans are suspension feeders that grow on substrates such as shells, rocks, algae and seagrasses and are prey to micro-predators, such as worms and small crustaceans and gastropods. As bio-constructors, they are known to increase the available habitat and overall diversity of marine ecosystems, making them an important component of many benthic communities. Nevertheless, bryozoans remain a relatively understudied phylum as they have no direct economic value and are generally small, cryptic and difficult to identify without the use of a microscope. Given their obscurity relative to more prominently studied taxa, the diversity of bryozoans around Galiano Island, as throughout the region, is likely under-reported.
Of the 17 bryozoan species reported for Galiano Island, one is introduced to British Columbia: Schizoporella japonica Ortmann, 1890 (orange ripple bryozoan) (Fig.

As aquatic suspension feeders, bryozoans pluck food particles from the surrounding water with a specialised structure called a lophophore. Each individual or zooid is typically protected within a covering of calcium carbonate or chitinous material. The protective features and structure of this covering are polymorphic and used to distinguish species. Bryozoan species and their geographic ranges are often listed in invertebrate identification keys (e.g.
Phylum
[1 class: 1 order: 1 family: 1 genus: 1 species]
‘Phoronida’ is the plural form of the phylum’s type genus Phoronis, likely derived from the Latin 'Phoronis' (the last name of the mythical Io, priestess of Argos).
Phoronida (horseshoe worms) are a small phylum of soft-bodied filter-feeding marine animals that form upright tubes of chiton to support their bodies. Worldwide the phylum includes two genera, comprising 13 recognised species (

Phoronids are benthic organisms that occur at depths ranging from the intertidal zone to about 400 m of depth in all oceans and seas throughout the world, except the Antarctic seas (
Phoronids are called “horseshoe worms” because the top of their worm-like body contains rows of ciliated tentacles forming a horseshoe shape. This is the lophophore—the feeding organ they share in common with the Brachiopoda, Bryozoa and Entoprocta. References to the phoronids reported for the region may be found in
Phylum
[5 classes: 14 orders: 20 families: 30 genera: 41 species]
‘Echinodermata’ is rooted in the Ancient Greek ἐχῖνος (echīnos), meaning “hedgehog”, and δέρμα (derma), “skin”.
Echinoderms are a phylum of marine invertebrates characterised by tough, spiny skin and radial symmetry, containing over 7,000 recognised species worldwide (
Echinoderms, especially sea stars, are an ecologically important component of the marine fauna of the Northeast Pacific. The term “keystone species” was first coined by

Under-represented echinoderms in the Galiano Island record include the holothuroids and ophiuroids. This gap is likely due to sampling bias, as many of these taxa are found at deeper depths than have been sampled locally or otherwise tend to be buried beneath rock and sediment. Gaps otherwise relate to limits in taxonomic knowledge. For example, the genus Henricia Gray, 1840 is highly variable and in need of clarification. Genetic studies are underway to sort out many suspected new species of Henricia between Alaska and California, led by the efforts of Roger Clark, Doug Eernisse, Megumi Strathmann and Christopher Mah. Leptasterias hexactis (Stimpson, 1862) also belongs to a species complex (Leptasterias aequalis species complex) requiring further study. Regional accounts of echinoderms include
Subphylum
[1 class: 3 orders: 12 families: 24 genera: 33 species]
The name ‘Tunicata’ comes from the Latin tunicatus, past particle of the verb tunicare, “to clothe with a tunic.”
Tunicates are filter-feeding marine animals that derive their name from their protective exoskeleton, called the tunic, which is formed of proteins and carbohydrates. A subphylum of Chordata, Tunicata are traditionally divided into three classes—Ascidiacea, Thaliacea and Appendicularia—though recent evidence suggests this group is paraphyletic (
Ascidians are sessile organisms found at all ocean depths worldwide, attaching to natural surfaces such as rocky outcrops and invertebrate shells, as well as artificial structures such as pilings, docks and boat hulls. Notable reports for Galiano Island include several non-native species that have a long history of invasion, often resulting in measurable impacts in other parts of Canada and around the world (e.g.

Tunicates are generally under-represented in this study as they are easily overlooked; new species are still being discovered in British Columbia (
Identification of tunicates is often based on gross morphological features, though dissection is occasionally required. Ascidians may be categorised as solitary, social or colonial according to variations on a basic body plan. The individuals, called zooids, are very small (measuring a few mm), but the colonies can be up to several metres large. Though tunicates are generally easily overlooked, often confused with other groups and challenging to identify to species, there are excellent resources available to support their identification, including: popular field guides (
Class
[11 orders: 29 families: 58 genera: 78 species]
Both Actinopterygii (ray-finned fish) and Chondrichthyes (cartilaginous fish) are treated below.
‘Actinopterygii’ is formed from the Latin 'actino-', “having rays”, and the Ancient Greek πτέρυξ (ptérux), meaning “wings” or “fins.” ‘Chondrichthyes’ comes from the Greek χόνδρος (khóndros), meaning "cartilage", and ἰχθύς (ikhthū́s), meaning "fish".
Actinopterygii contain over 33,000 described taxa and Chondrichthyes (cartilaginous fish) over 1,100 taxa, including both freshwater and marine species (
Marine fishes dwell in various habitats, from deep sea (pelagic) to near-shore and intertidal environments. Taxa under-represented in this dataset include chondrichthyes (cartilaginous fish), myxinids (hagfish) and osmerids (smelts). These gaps reflect search efforts mostly biased toward rocky reefs and exposed beach locales, to waters < 30 m in depth. Agonids (poachers), pleuronectids (right-eyed flounders) and zoarcids (eelpouts) are also under-represented. These are primarily subtidal, soft substrate inhabitants, indicating that sampling has been limited in these habitats.
Rockfish, Sebastes G. Cuvier, 1829, are a long-lived and diverse genus of fish in the Northeast Pacific, 11 of which are represented in the Galiano Island record. Species present include the threatened Sebastes maliger (Jordan & Gilbert, 1880) (quillback rockfish) and S. ruberrimus (Cramer, 1895) (yellow-eye rockfish) (Fig.

Squalus suckleyi Girard, 1854 (North Pacific spiny dogfish), reported for Galiano Island, is a species of special concern in Canada (
Class
[3 orders: 3 families: 3 genera: 4 species]
Chondrichthyes are discussed above, alongside Actinopterygii. For annotated records of cartilaginous fishes reported for Galiano Island, see supplementary materials (Suppl. material
Class
[2 orders: 7 families: 12 genera: 13 species]
The name ‘Mammalia’ was coined in 1758 by Linnaeus, from Late Latin (neuter plural) 'mammalis', meaning “of the breast.”
Mammalia are a class of vertebrates containing 6,400 species worldwide (
Among locally occurring cetaceans are: baleen whales, distinguished by their enlarged head containing baleen plates suspended from the upper jaw, their double blowhole and their thick blubber; and toothed whales, distinguished by their teeth and single external blowhole. Certain cetaceans, such as Balaenoptera physalus (Linnaeus, 1758) (fin whale) and Balaenoptera acutorostrata Lacépède, 1804 (common minke whale) have been recorded infrequently in local waters, whereas others, such as Megaptera novaeangliae Borowski, 1781 (humpback whale) and Phocoena phocoeana (Linnaeus, 1758) (harbour porpoise), are commonly seen. Among carnivores, both aquatic pinnipeds, such as Phoca vitulina richardii Gray, 1864 (harbour seal) and semi-aquatic fissipeds, such as Lontra canadensis (Schreber, 1777) (river otter), are represented. Presently, there are no known haulouts of Mirounga angustirostris (Gill, 1866) (northern elephant seal) around Galiano Island, though they are known not far away on Salt Spring Island.
Populations of Eschrichtius robustus (Lilljeborg, 1861) (grey whale), M. novaeangliae, P. phocoena and the southern resident population of Orcinus orca (Linnaeus, 1758) (orca) (Fig.

Discussion
Extensive efforts are being made to promote the availability of biodiversity data, which have increased considerably over the course of the last decade (
While there is growing recognition of the value of involving citizen scientists in biodiversity research (
Within the Salish Sea, PMLS records currently account for ~ 248,000 observations from > 4,800 dives, documenting > 1,100 species. These records have supported numerous studies, providing information for a broad range of taxonomic and ecological research (e.g.
Our synthesis sheds light on the complementarity of various types of sampling effort, pointing to ways in which improved data sharing and collaboration might enable more systematic biodiversity research practices. These lessons derive from a basic realisation: different types of sampling effort have distinct constraints in terms of taxonomic, spatial and temporal resolution, but when combined, can contribute to more robust biodiversity baselines. Below, we discuss the limitations and complementarity of the various data sources synthesised in this study.
Collections
Collections are of tremendous value for taxonomic research, not only providing the oldest records in this study, but also yielding the highest ratio of novel species reports to occurrence records (1:4) as compared to crowd-sourced observations (1:38), ecological surveys (1:66) and PMLS records (1:67). Voucher specimens are relatively few in this study, with low spatial coverage, yet offer improved taxonomic resolution of difficult groups such as molluscs, crustaceans and annelids. Specimens are necessary to identify many taxa, which often entails microscopy, dissection and genetic study. Yet, processing specimens and making such data available is notoriously slow. Worldwide, it is estimated that only 10% of biospecimen data are available in digital form (
Crowd-sourced data
Observations on iNaturalist and the BC Cetacean Sightings Network (BCCSN) have greater spatial coverage than other sources included in this dataset, yet are more diffuse than systematic efforts such as the PMLS. In this study, data from the PMLS and research collections largely predate observations derived from these platforms. That crowd-sourced observations nevertheless contribute 18% of novel species reports in this study demonstrates the value of these data, which may supplement baseline datasets, even in areas historically subject to intensive search efforts. In contrast to PMLS observations, which are limited from the subtidal to a depth of 30 m, these records largely represent casual intertidal beach-combing and nearshore cetacean observations. Exceptions include iNaturalist observations yielded by SCUBA and snorkelling and those posted from research programmes, such as those of
Ecological studies
Ecological studies are more narrowly focused than other sampling approaches represented in this dataset. These studies have limited temporal and spatial resolution and focus on: a) anadromous salmonids (
Pacific Marine Life Surveys (PMLS)
Ongoing since 1967, the PMLS are systematic marine biodiversity surveys, based on the roving diver methodology (
Conclusion: Strengthening connections in the biodiversity research community
Our study shows the potential for communities to synthesise comprehensive baseline datasets, which may form the basis for more coordinated and systematic biodiversity monitoring efforts. To realise this potential, however, the global community has been challenged to rethink the dynamics of biodiversity research practice (
This study sets an example for a community science initiative that both contributes to the global biodiversity research community and actively uses its own data to further its research objectives in collaboration with research scientists. We join
Depending on the data available from place to place, systematic community-based sampling efforts, as exemplified by the PMLS in this study, may prove critical in the establishment of robust biodiversity baselines. When combined with citizen science observations, ecological data and research collections, these surveys can also bolster the data available through major aggregators, such as the Global Biodiversity Information Facility (GBIF). The open-source biodiversity informatics framework that we are developing harnesses readily accessible tools such as Google Sheets, GitHub and JavaScript code to facilitate curation processes that integrate data available through iNaturalist and GBIF with other sources of local biodiversity knowledge. This inclusive data management system will enable communities to synthesise data from a diversity of sources, establishing baselines for localised biodiversity monitoring efforts. Scaled up, this system can support distributed biodiversity monitoring networks that synthesise data generated by communities throughout regions such as the Salish Sea. Our study represents a gesture toward the development of such a framework at a time of critical need.
Acknowledgements
This research was supported by the Institute for Multidisciplinary Ecological Research in the Salish Sea and the Ian McTaggart Cowan Professorship at the University of Victoria School of Environmental Studies. The authors thank the following individuals for their assistance and review of this work: Sheila Byers, Jim Carlton, Hannah Carpendale, Alice Chang, Henry Choong, Roger Clark, Keith Erickson, Aquila Flower, Trevor Goward, Doug Ernisse, Stefania Gorgopa, Bernie Hanby, Gregory Jensen, Gretchen Lambert, Sandra Lindstrom, Milton Love, Jeff Marliave, Claudia Mills, Randal Mindell, Bruce Ott, Wyatt Patry, Amy Rowley, Linda Schroeder, Kathryn Sobocinski, Karolle Wall, Matt Whalen, Mary Wicksten and Gary Williams. We also remember the late Bill Austin and Henry Reiswig, who were at the forefront of research focusing on Porifera. Since Bill and Henry’s passing, Bruce Ott has carried on regional taxonomic efforts describing these obscure organisms.
References
- American seashells.2nd ed.Van Nostrand Reinhold Company,New York,541pp.
- Contributions to the Natural History of the United States of America: pt. 3. Discophoræ. pt. 4. Hydroidæ. pt. 5. Homologies of the Radiata.Little, Brown and Company,Boston,446pp.
- Evidence that microorganisms at the animal-water interface drive sea star wasting disease.Frontiers in Microbiology11:3278.
- Pressurized seawater as an antifouling treatment against the colonial tunicates Botrylloides violaceus and Botryllus schlosseri in mussel aquaculture.Aquatic Invasions6(4):465‑476. https://doi.org/10.3391/ai.2011.6.4.12
- Temporal and spatial development of an infestation of Styela clava on mussel farms in Malpeque Bay, Prince Edward Island, Canada.Aquatic Invasions4(1):189‑194. https://doi.org/10.3391/ai.2009.4.1.19
- Two new species in the family Axinellidae (Porifera, Demospongiae) from British Columbia and adjacent waters.ZooKeys338:11. https://doi.org/10.3897/zookeys.338.5535
- Taxonomic review of Hadromerida (Porifera, Demospongiae) from British Columbia, Canada, and adjacent waters, with the description of nine new species.Plazi.org taxonomic treatments databasehttps://doi.org/10.15468/w8macu
- Systematic list of amphipods found in British Columbia. https://ibis.geog.ubc.ca/biodiversity/efauna/documents/AmphibodsofBCBaldwin09.pdf. Accessed on: 2021-2-13.
- Chitons of British Columbia. https://ibis.geog.ubc.ca/biodiversity/efauna/documents/ChitonsofBCBaldwin2009.pdf. Accessed on: 2021-2-06.
- Polychaete worms of British Columbia. https://ibis.geog.ubc.ca/biodiversity/efauna/documents/PolychaetesofBCBaldwin2009.pdf. Accessed on: 2021-2-09.
- Checklist of the cnidarian medusae (jellyfish) and ctenophorans (comb jellies) of British Columbia. https://ibis.geog.ubc.ca/biodiversity/efauna/documents/JellyfishofBC0409.pdf. Accessed on: 2021-2-13.
- The lophophorate phyla of British Columbia: entoprocts, bryozoans, phoronids, and brachiopods. https://ibis.geog.ubc.ca/biodiversity/efauna/documents/Lophophorates%20of%20BCBaldwin2009.pdf. Accessed on: 2021-2-13.
- Checklist of the invertebrate chordata and the hemichordata of British Columbia (tunicates and acorn worms). https://ibis.geog.ubc.ca/biodiversity/efauna/documents/TunicatesInvertebrateChordatesofBCBaldwin2009.pdf. Accessed on: 2021-2-13.
- The nemerteans (ribbon worms) of British Columbia. https://ibis.geog.ubc.ca/biodiversity/efauna/documents/NemerteansofBCJuly2010Baldwin.pdf. Accessed on: 2021-2-05.
- Checklist of the tapeworms, flukes, and monogeneans of British Columbia. https://ibis.geog.ubc.ca/biodiversity/efauna/documents/RibbonWormsPlatyhelminthesBaldwinJuly2010.pdf. Accessed on: 2021-2-05.
- Checklist of the shrimps, crabs, lobsters and crayfish of British Columbia 2011 (Order Decapoda). https://ibis.geog.ubc.ca/biodiversity/efauna/documents/DecapodsofBCBaldwin2011.pdf. Accessed on: 2021-2-13.
- Checklist of the barnacles of British Columbia. https://ibis.geog.ubc.ca/biodiversity/efauna/documents/BarnaclesofBCBaldwin2016.pdf. Accessed on: 2021-2-13.
- Research applications of primary biodiversity databases in the digital age.PLOS One14(9):e0215794. https://doi.org/10.1371/journal.pone.0215794
- Sabellidae (Polychaeta) principally from the northeast Pacific Ocean.Journal of the Fisheries Board of Canada36(8):869‑882. https://doi.org/10.1139/f79-125
- On some Cossuridae and Maldanidae (Polychaeta) from Washington and British Columbia.Canadian Journal of Fisheries and Aquatic Sciences38(6):633‑637. https://doi.org/10.1139/f81-084
- BC Ecosystem Explorer. https://a100.gov.bc.ca/pub/eswp/. Accessed on: 2021-1-23.
- Pacific coast nudibranchs, supplement II: new species to the Pacific Coast and new information on the oldies.Proceedings of the California Academy of Sciences55(2):11‑54.
- Eastern Pacific nudibranchs.Sea Challengers,Monterey,137pp.
- A checklist of Polychaeta recorded from British Columbia since 1923, with references to name changes, descriptions, and synonymies. I. Errantia.Canadian Journal of Zoology45(6):1049‑1059. https://doi.org/10.1139/z67-114
- A checklist of Polychaeta recorded from British Columbia since 1923, with references to name changes, descriptions, and synonymies. II. Sedentaria.Canadian Journal of Zoology46(3):557‑567. https://doi.org/10.1139/z68-073
- Annelida. Polychaeta Errantia. Canadian Pacific Fauna.University of Toronto Press for the Fisheries Research Board of Canada,Toronto,100pp.
- Annelida. Polychaeta Sedentaria.University of Toronto Press for the Fisheries Research Board of Canada,Toronto,139pp.
- Polychaeta from British Columbia; with a note on some western Canadian arctic forms.Canadian Journal of Zoology40(4):571‑577. https://doi.org/10.1139/z62-050
- Prodrome for a distributional check-list and bibliography of the recent marine mollusca of the west coast of Canada.Fisheries Research Board of CanadaTech Report 2:261 pp.
- The living Brachiopoda of British Columbia.Syesis5:73‑82.
- The distribution of the planktonic chaetognatha in the Pacific and their relationship to the water masses.Limnology and Oceanography4(1):1‑28. https://doi.org/10.4319/lo.1959.4.1.0001
- Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel: the Annelida. Part 1. Oligochaeta and Polychaeta: Phyllodocida (Phyllodocidae to Paralacydoniidae).US Department of the Interior, Minerals Management Service, Pacific OCS Region (No. 42).406 pp.
- Family Pholadidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 5. The Annelida Part 2. Polychaeta: Phyllodocida (Syllidae and Scale-bearing Families), Amphinomida, and Eunicida.Santa Barbara Museum of Natural History,Santa Barbara,378pp.
- Family Orbiniidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 6. The Annelida Part 3. Polychaeta: Orbiniidae to Cossuridae.Santa Barbara Museum of Natural History,Santa Barbara,377pp.
- Family Magelonidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 6. The Annelida Part 3. Polychaeta: Orbiniidae to Cossuridae.Santa Barbara Museum of Natural History,Santa Barbara,377pp.
- Family Cirratulidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 6. The Annelida Part 3. Polychaeta: Orbiniidae to Cossuridae.Santa Barbara Museum of Natural History,Santa Barbara,377pp.
- Family Paraonidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 6. The Annelida Part 3. Polychaeta: Orbiniidae to Cossuridae.Santa Barbara Museum of Natural History,Santa Barbara,377pp.
- Family Spionidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 6. The Annelida Part 3. Polychaeta: Orbiniidae to Cossuridae.Santa Barbara Museum of Natural History,Santa Barbara,377pp.
- Family Phyllodocidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 4. The Annelida Part 1. Oligochaeta and Polychaeta: Phyllodocida (Phyllodocidae to Paralacydoniiae).Santa Barbara Museum of Natural History,Santa Barbara,406pp.
- Family Capitellidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 7. The Annelida Part 4. Polychaeta: Flabelligeridae to Sternapsidae.Santa Barbara Museum of Natural History,Santa Barbara,348pp.
- Family Flabelligeridae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 7. The Annelida Part 4. Polychaeta: Flabelligeridae to Sternapsidae.Santa Barbara Museum of Natural History,Santa Barbara,348pp.
- Family Opheliidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 7. The Annelida Part 4. Polychaeta: Flabelligeridae to Sternapsidae.Santa Barbara Museum of Natural History,Santa Barbara,348pp.
- Family Oweniidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 7. The Annelida Part 4. Polychaeta: Flabelligeridae to Sternapsidae.Santa Barbara Museum of Natural History,Santa Barbara,348pp.
- Family Scalibregmatidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 7. The Annelida Part 4. Polychaeta: Flabelligeridae to Sternapsidae.Santa Barbara Museum of Natural History,Santa Barbara,348pp.
- Phylum Bryozoa Ehrenberg, 1831.Zootaxa3703:67‑74. https://doi.org/10.11646/zootaxa.3703.1.14
- Corals of British Columbia. https://ibis.geog.ubc.ca/biodiversity/efauna/documents/CoralChecklist2011BoutillierGillespie.pdf. Accessed on: 2021-2-04.
- Sipuncula: an emerging model of spiralian development and evolution.The International Journal of Developmental Biology58:485‑499. https://doi.org/10.1387/ijdb.140095mb
- Seasonality of hydroids (Hydrozoa, Cnidaria) from an intertidal pool and adjacent subtidal habitatas at Race Rocks, off Vancouver Island, Canada.Scientia Marina60(1):89‑97.
- Further notes on the Leptolida (Hydrozoa: Cnidaria) from Canadian Pacific Waters.Leiden Zoologische verhandelingen323(31):37‑68.
- How many species of mammals are there?Journal of Mammalogy99(1):1‑14. https://doi.org/10.1093/jmammal/gyx147
- A guide to rockfishes, thornyheads and scorpionfishes of the Northeast Pacific.University of California Press,Berkeley,185pp.
- Common and scientific names of aquatic invertebrates from the United States and Canada: Cnidaria and Ctenophora. 2nd ed.Special Publication 28.American Fisheries Society,Bethesda,115pp. https://doi.org/10.47886/9781888569643
- Similiclava nivea (Cnidaria: Hydrozoa: Similiclavidae): a new family, genus and species of athecate hydroid from the Pacific coast of North America.Journal of Natural History49:735‑753. https://doi.org/10.1080/00222933.2014.979261
- Names of hydroids (Cnidaria, Hydrozoa) established by Charles McLean Fraser (1872–1946), excluding those from Allan Hancock Expeditions.Zootaxa4487(1). https://doi.org/10.11646/zootaxa.4487.1.1
- A Stó: lo-Coast Salish historical atlas/edited by Keith Thor Carlson.University of Washington Prress,Seattle,192pp.
- The Light and Smith Manual, intertidal invertebrates from Central California to Oregon.4th ed.University of California Press,Berkeley,1019pp. https://doi.org/10.1525/9780520930438
- A tri-oceanic perspective: DNA barcoding reveals geographic structure and cryptic diversity in Canadian polychaetes.PLOS One6(7). https://doi.org/10.1371/journal.pone.0022232
- Contribution of citizen science towards international biodiversity monitoring.Biological Conservation213:280‑294. https://doi.org/10.1016/j.biocon.2016.09.004
- Principles and methods of data cleaning: primary species and species-occurrence data. version 1.0. Report for the Global Biodiversity Information Facility.Global Biodiversity Information Facility,Copenhagen.
- Hydroids of the genus Sertularella (Cnidaria: Hydrozoa: Sertulariidae) from the Pacific coast of Canada in the collection of the Royal Ontario Museum, with descriptions of four new species.Zootaxa3925(3):387. https://doi.org/10.11646/zootaxa.3925.1
- High resolution mapping of community structure in three glass sponge reefs (Porifera, Hexactinellida).Marine Ecology Progress Series417:97‑113. https://doi.org/10.7939/r36k3q
- The dorid nudibranchs Peltodoris lentiginosa and Archidoris odhneri as predators of glass sponges.Invertebrate Biology131(2):75‑81. https://doi.org/10.1111/j.1744-7410.2012.00262.x
- The shallow water chiton fauna of the Salish Sea.Festivus52(4):294‑315. https://doi.org/10.54173/F524294
- Fishes of the Pacific coast of Canada. Bulletin No. 68.2nd ed.Fisheries Research Board of Canada,Ottawa,443pp.
- Bivalve seashells of western North America: Marine bivalve mollusks from Arctic Alaska to Baja California.Santa Barbara Museum of Natural History,Santa Barbara,764pp.
- Papers from the Harriman Alaska Expedition, 20. The nemerteans of the expedition.Proceedings of the Washington Academy of Sciences3:1‑110.
- Nemerteans of the Pacific coast of North America, part 2.Harriman Alaska Expedition11:11‑220.
- Nemerteans of the west and northwest coast of America.Bulletin of the Museum of Comparative Zoology. Harvard.47:1‑319.
- Geographical distributions of the nemerteans of the Pacific coast of North America, with descriptions of two new species.Journal of the Washington Academy of Sciences34:27‑32.
- Platyhelminthes.Current Biology27(7). https://doi.org/10.1016/j.cub.2017.02.016
- The barnacles of British Columbia. British Columbia Provincial Handbook No. 7. Royal British Columbia Museum, Victoria 71pp.
- COSEWIC assessment and status report on the North Pacific Spiny Dogfish Squalus suckleyi in Canada.Committee on the Status of Endangered Wildlife in Canada. URL: https://publications.gc.ca/collections/collection_2012/ec/CW69-14-644-2012-eng.pdf
- Cephalopods (octopuses and squids) of British Columbia. https://ibis.geog.ubc.ca/biodiversity/efauna/documents/OctopusesSquidsofBC.pdf. Accessed on: 2021-2-06.
- Super suckers: the giant Pacific octopus and other cephalopods of the Pacific coast.Harbour Publishing,Madeira Park,256pp.
- A key to the Mysidacea of the Pacific Northwest.Canadian Journal of Zoology64(6):1201‑1210. https://doi.org/10.1139/z86-179
- A comparative analysis of biodiversity and distribution of shallow-water marine isopods (Crustacea: Isopoda) from polar and temperate waters in the East Pacific.Belgian Journal of Zoology136(2):219.
- A phylogenomic framework and timescale for comparative studies of tunicates.BMC Biology16(1):39. https://doi.org/10.1186/s12915-018-0499-2
- Sharks of the Pacific Northwest, including Oregon, Washington, British Columbia & Alaska.Harbour Publishing,Madeira Park,144pp.
- World Porifera Database. http://www.marinespecies.org/porifera. Accessed on: 2021-12-22.
- Assessing condition and ecological role of deep-water biogenic habitats: glass sponge reefs in the Salish Sea.Marine Environmental Research141:88‑99. https://doi.org/10.1016/j.marenvres.2018.08.002
- The hidden biology of sponges and ctenophores.Trends in Ecology & Evolution30(5):282‑291. https://doi.org/10.1016/j.tree.2015.03.003
- Foundation Species, Non-trophic Interactions, and the Value of Being Common.iScience13:254‑268. https://doi.org/10.1016/j.isci.2019.02.020
- Phylum: Phoronida. In:Grzimek's Animal Life Encyclopedia. 2: Protostomes.2nd ed.Thompson Gale,9000pp.
- 1999 / 2000 urban salmon habitat program final report.Galiano Conservancy Association, Galiano Island, BC.Unpublished report.
- Recovery strategy for the transient killer whale (Orcinus orca) in Canada. Species at Risk Act recovery strategy series.Fisheries and Oceans Canada.
- Management plan for the Pacific harbour porpoise (Phocoena phocoena) in Canada. Species at Risk Act management plan series.Fisheries and Oceans Canada.
- Management plan for the offshore killer whale (Orcinus orca) in Canada. Species at Risk Act management plan series.Fisheries and Oceans Canada.
- Management plan for the Eastern Pacific grey whale (Eschrichtius robustus) in Canada. Species at Risk Act management plan series.Fisheries and Oceans Canada.
- Recovery strategy for the North Pacific humpback whale (Megaptera novaeangliae) in Canada. Species at Risk Act recovery strategy series.Fisheries and Oceans Canada.
- Recovery strategy for the northern and southern resident killer whales (Orcinus orca) in Canada. Species at Risk Act recovery strategy series.Fisheries & Oceans Canada.
-
Management Plan for the Yelloweye Rockfish (Sebastes ruberrimus) in Canada [Proposed]. Species at Risk Act Management Plan Series. Fisheries and Oceans Canada, Ottawa. iv + 32 pp.
- Rockfish conservation areas. https://www.pac.dfo-mpo.gc.ca/fm-gp/maps-cartes/rca-acs/index-eng.html. Accessed on: 2021-2-18.
- Marine mammals of British Columbia.Royal British Columbia Museum,Victoria (BC, Canada),464pp.
- List of bivalve molluscs from British Columbia, Canada. https://ibis.geog.ubc.ca/biodiversity/efauna/documents/Bivalve_List_11Oct2013.pdf
- Intertidal bivalves: a guide to the common marine bivalves of Alaska.University of Alaska Press,Fairbanks,152pp.
- Hydroids of the Pacific coast of Canada and the United States.University of Toronto Press,Toronto,207pp. https://doi.org/10.3138/9781487595548
- First confirmed records of the non-native amphipod Caprella mutica (Schurin 1935) along the coast of British Columbia, Canada, and the potential for secondary spread via hull fouling.Aquatic Invasions4(3):495‑499. https://doi.org/10.3391/ai.2009.4.3.10
- Temporal variations in the carotenoid composition and content of Euphausia pacifica Hansen in Saanich Inlet, British Columbia.Journal of Experimental Marine Biology and Ecology148(1):93‑104. https://doi.org/10.1016/0022-0981(91)90149-Q
- Patterns in the distribution and abundance of selected zooplankton species from the coast of British Columbia.Biological Oceanography1(3):255‑270.
- British Columbia pelagic marine Copepoda: an identification manual and annotated bibliography. Canadian Special Publication of Fisheries and Aquatic Sciences.62.Department of Fisheries and Oceans, 1979,Ottawa,53pp.
- BC tunicates. http://taxonomy.royalbcmuseum.bc.ca/Taxonomy/viewer/BC_Tunicates.aspx. Accessed on: 2021-2-13.
- Non-indigenous invertebrate species in the marine fouling communities of British Columbia, Canada.BioInvasions Records5(4):205‑212. https://doi.org/10.3391/bir.2016.5.4.03
- No longer Demospongiae: Homoscleromorpha formal nomination as a fourth class of Porifera.Hydrobiologia687(1):3‑10. https://doi.org/10.1007/s10750-011-0842-x
- GBIF occurrence download. https://doi.org/10.15468/dl.c2dd4a. Accessed on: 2021-2-18.
- Pacific coast subtidal marine invertebrates.Sea Challengers,Monterey,107pp.
- Pycnopodia helianthoides.IUCN Red List of Threatened Specieshttps://doi.org/10.2305/iucn.uk.2021-1.rlts.t178290276a197818455.en
- Cladocera of British Columbia. https://ibis.geog.ubc.ca/biodiversity/efauna/documents/CladoceraChecklistEFaunaBCGardiner20190103.pdf. Accessed on: 2021-2-13.
- Landscape construction, ownership and social change in the Southern Gulf Islands of British Columbia.Canadian Journal of Archaeology/Journal Canadien d'Archéologie38(1):211‑249.
- Terraforming and monumentality as long-term social practice in the Salish Sea region of the Northwest Coast of North America.Hunter Gatherer Research3(1):107‑132. https://doi.org/10.3828/hgr.2017.7
- ‘Fish’ (Actinopterygii and Elasmobranchii) diversification patterns through deep time.Biological Reviews91(4):950‑981. https://doi.org/10.1111/brv.12203
- Shells & shellfish of the Pacific Northwest.Harbor Publishing,Madeira Park,272pp.
- Intertidal clam resources (manila, littleneck and butter clam) volume II: the southern inside waters of Vancouver Island and the British Columbia mainland. Canadian manuscript report of fisheries and aquatic sciences 2417.Fisheries and Oceans Canada, Ottawa, 245 pp.
- Whelks to whales.2nd ed.Harbour Publishing,Maideira Park,328pp.
- Some Cumacea of the Vancouver Island region.Contributions to Canadian Biology and Fisheries6(1):23‑40. https://doi.org/10.1139/f31-003
- Pacific fishes of Canada.180.Bulletin of the Fisheries Research Board of Canada,Ottawa,740pp.
- Quantifying marine mammal hotspots in British Columbia, Canada.Ecosphere8(7):e01884. https://doi.org/10.1002/ecs2.1884
- Mollusca (Molluscs).eLShttps://doi.org/10.1038/npg.els.0001598
- Family Onuphidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 4. The Annelida Part 2. Polychaeta: Phyllodocida (Syllidae and Scale-bearing Families), Amphinomida, and Eunicida.Santa Barbara Museum of Natural History,Santa Barbara,378pp.
- Family Lumbrineridae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 4. The Annelida Part 2. Polychaeta: Phyllodocida (Syllidae and Scale-bearing Families), Amphinomida, and Eunicida.Santa Barbara Museum of Natural History,Santa Barbara,378pp.
- Family Glyceridae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 4. The Annelida Part 1. Oligochaeta and Polychaeta: Phyllodocida (Phyllodocidae to Paralacydoniiae).Santa Barbara Museum of Natural History,Santa Barbara,369pp.
- Family Goniadidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 4. The Annelida Part 1. Oligochaeta and Polychaeta: Phyllodocida (Phyllodocidae to Paralacydoniiae).Santa Barbara Museum of Natural History,Santa Barbara,369pp.
- Family Nephtyidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 4. The Annelida Part 1. Oligochaeta and Polychaeta: Phyllodocida (Phyllodocidae to Paralacydoniiae).Santa Barbara Museum of Natural History,Santa Barbara,369pp.
- Family Ampharetidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 7. The Annelida Part 4. Polychaeta: Flabelligeridae to Sternapsidae.Santa Barbara Museum of Natural History,Santa Barbara,406pp.
- Family Terebellidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 7. The Annelida Part 4. Polychaeta: Flabelligeridae to Sternapsidae.Santa Barbara Museum of Natural History,Santa Barbara,406pp.
- Family Trichobranchidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 7. The Annelida Part 4. Polychaeta: Flabelligeridae to Sternapsidae.Santa Barbara Museum of Natural History,Santa Barbara,406pp.
- Sedentariate and archiannelid polychaetes of British Columbia and Washington.Canadian Bulletin of Fisheries and Aquatic Sciences209:1‑144.
- Brachiopoda. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel. Santa Barbara Museum of Natural History, vol. 4.Santa Barbara Museum of Natural History,Santa Barbara,305pp.
- Systema Porifera: a guide to the classification of sponges.Springer US,New York,XCIV, 1707pp. https://doi.org/10.1007/978-1-4615-0747-5
- DNA barcoding reveals cryptic diversity in the peanut worm Sipunculus nudus.Molecular Ecology Resources13(4):596‑606. https://doi.org/10.1111/1755-0998.12097
- Compendium of bivalves: A full-color guide to 3,300 of the world's marine bivalves.Conchbooks,Hackenheim,904pp.
- Three New Species Of Lebbeus (Crustacea: Decapoda: Hippolytidae) from the Northeastern Pacific.Zenodohttps://doi.org/10.5281/zenodo.175047
- Crabs and shrimps of the Pacific coast, a guide to shallow-water decapods from Southeastern Alaska to the Mexican border.MolaMarine,Bremerton,240pp.
- Beneath Pacific tides: subtidal invertebrates of the West Coast.MolaMarine,Bremerton,296pp.
- Phylum Phoronida. In:Atlas of marine invertebrate larvae.Academic Press,Amsterdam, Netherlands,630pp.
- The basal unconformity of the Nanaimo Group, southwestern British Columbia: a Late Cretaceous storm-swept rocky shoreline.Canadian Journal of Earth Sciences43(8):1165‑1181. https://doi.org/10.1139/e06-046
- Salish Sea response to global climate change, sea level rise, and future nutrient loads.Journal of Geophysical Research: Oceans124(6):3876‑3904. https://doi.org/10.1029/2018jc014670
- Establishment of an interdisciplinary project to evaluate ecological implications of climate change in dry south coast forest ecosystems.Ministry of Forests, Lands and Natural Resource Operations.(Technical Report 086). URL: https://www.for.gov.bc.ca/hfd/pubs/docs/tr/TR086.pdf
- The distribution of the orange‐striped green anemone, Diadumene lineata, in relation to environmental factors along coastal British Columbia, Canada.Invertebrate Biology138(4). https://doi.org/10.1111/ivb.12268
- The genus Zelentia is an amphi-boreal taxon expanded to include three new species from the North Pacific and Atlantic oceans (Gastropoda: Nudibranchia: Trinchesiidae).Zootaxa4482(2):297‑321. https://doi.org/10.11646/zootaxa.4482.2.4
- The Emperor’s Cadlina, hidden diversity and gill cavity evolution: new insights for the taxonomy and phylogeny of dorid nudibranchs (Mollusca: Gastropoda).Zoological Journal of the Linnean Society189(3):762‑827. https://doi.org/10.1093/zoolinnean/zlz126
- Marine invertebrates of the Pacific northwest.2nd ed.University of Washington Press,Seattle,552pp.
- Southern Canada’s crisis ecoregions: identifying the most significant and threatened places for biodiversity conservation.Biodiversity and Conservation29(13):3573‑3590. https://doi.org/10.1007/s10531-020-02038-x
- Family Syllidae. In:Taxonomic atlas of the benthic fauna of the Santa Maria Basin and Western Santa Barbara Channel, vol. 4. The Annelida Part 2. Polychaeta: Phyllodocida (Syllidae and Scale-bearing Families), Amphinomida, and Eunicida.Santa Barbara Museum of Natural History,Santa Barbara,378pp.
- A synopsis and key to the recent Ophiuroidea of Washington State and southern British Columbia.Journal of the Fisheries Research Board of Canada26:1727‑1741. https://doi.org/10.1139/f69-160
- Marine life of the Pacific Northwest: a photographic encyclopedia of invertebrates, seaweeds and selected fishes.Harbour Publishing,Maideira Park,398pp.
- Coastal fishes of the Pacific northwest. Revised and expanded second edition.Harbour Publishing,Madeira Park,335pp.
- Distaplia alaskensis sp. nov. (Ascidiacea, Aplousobranchia) and other new ascidian records from south-central Alaska, with a redescription of Ascidia columbiana (Huntsman, 1912).Canadian Journal of Zoology79:1766‑1781. https://doi.org/10.1139/z01-141
- New records of ascidians from the NE Pacific: a new species of Trididemnum, range extension and redescription of Aplidiopsis pannosum (Ritter, 1899) including its larva, and several non-indigenous species.Zoosystema25(4):665‑679.
- Invasive sea squirts: a growing global problem.Journal of Experimental Marine Biology and Ecology342(1):3‑4. https://doi.org/10.1016/j.jembe.2006.10.009
- Adventures of a sea squirt sleuth: unraveling the identity of Didemnum vexillum, a global ascidian invader.Aquatic Invasions4(1):5‑28. https://doi.org/10.3391/ai.2009.4.1.2
- The Ascidiacea collected during the 2017 British Columbia Hakai MarineGEO BioBlitz.Zootaxa4657(3):401‑436. https://doi.org/10.11646/zootaxa.4657.3.1
- Sea cucumbers of British Columbia, Puget Sound and Southeast Alaska.Royal BC Museum and University of BC Press,Vancouver,176pp.
- Sea stars of British Columbia, Southeast Alaska and Puget Sound.UBC Press,Vancouver,186pp.
- Brittle stars, sea urchins and feather stars of British Columbia, southeast Alaska and Puget Sound. RBCM Handbook.Royal British Columbia Museum,Victoria,176pp.
- Deep-sea Echinodermata of British Columbia, Canada.Canadian Technical Report of Fisheries and Aquatic Sciences2929:1‑143.
- ‘No Personal Motive?’ Volunteers, biodiversity, and the false dichotomies of participation.Ethics, Place & Environment9(3):279‑298. https://doi.org/10.1080/13668790600893319
- The chaetognaths of western Canadian coastal waters.Journal of the Fisheries Research Board Canada12:593‑617.
- Indigenous marine resource management on the Northwest Coast of North America.Ecological Processes2(1). https://doi.org/10.1186/2192-1709-2-12
- Zooplankton communities in the Strait of Georgia, British Columbia, track large-scale climate forcing over the Pacific Ocean.Progress in Oceanography115:90‑102. https://doi.org/10.1016/j.pocean.2013.05.025
- Resource inventory of marine and estuarine fishes of the west coast and Alaska: a checklist of North American Pacific and Arctic Ocean species from Baja California to the Alaska-Yukon border.Seattle, Washington: U.S. Department of the Interior, U.S. Geological Survey, Biological Resources Division.OCS Study MMS 2005-030 and USGS/NBII 2005-001.
- Certainly more than you want to know about the fishes of the Pacific Coast.Really Big Press,Santa Barbara,650pp.
- Preliminary investigation on aquatic invasive species of marine and estuarine macrobenthic invertebrates on floating structures in five British Columbia harbours.Fisheries and Oceans Canada, Science Branch-Pacific Region, DFO-UBC Centre for Aquaculture and Environmental Research,Vancouver,30pp.
- Giant axons and escape swimming in Euplokamis dunlapae (Ctenophora: Cydippida).The Biological Bulletin182(2):248‑256. https://doi.org/10.2307/1542118
- Biodiversity stability of shallow marine benthos in strait of Georgia, British Columbia, Canada through climate regimes, overfishing and ocean acidification.Biodiversity Loss in a Changing Planethttps://doi.org/10.5772/24606
- Seabed biodiversity shifts identify climate regimes: The 2011 climate regime shift and associated cascades.Selected Studies in Biodiversityhttps://doi.org/10.5772/intechopen.71599
- 2011 climate regime: revealed by seabed biodiversity. In:State of the physical, biological and selected fishery resources of Pacific Canadian marine ecosystems in 2017. Canadian Technical Report of Fisheries and Aquatic Sciences.3266.Fisheries & Oceans Canada Pacific Biological Station,Nanaimo,245pp.
- Formation, persistence, and recovery of glass sponge Reefs: a case study.Invertebrates - Ecophysiology and Managementhttps://doi.org/10.5772/intechopen.82325
- Climate and weather factors affecting winter sheltering by shoreline Copper Rockfish Sebastes caurinus in Howe Sound, British Columbia.Scientific Reports10(1). https://doi.org/10.1038/s41598-020-71284-4
- Reconstructing lingcod biomass in Georgia Strait and the effect of marine reserves on lingcod populations in Howe Sound.University of British ColumbiaURL: https://open.library.ubc.ca/collections/ubctheses/831/items/1.0074836
- An updated classification of the recent Crustacea.Science Series 39.Natural History Museum of Los Angeles CountyURL: http://zoobank.org/646d0814-31c7-446d-9282-6d872d6850da
- Description, external morphology, and natural history observations of Nebalia hessleri, new species (Phyllocarida: Leptostraca), from southern California, with a key to the extant families and genera of the Leptostraca.Journal of Crustacean Biology16(2):347‑372. https://doi.org/10.1163/193724096X00153
- An updated classification of the recent Crustacea.Natural History Museum of Los Angeles County Contributions in Science39:495-497.
- 37. Note on the systematic position and distribution of the actinian Sagartia luciœ.Proceedings of the Zoological Society of London91(4):729‑739. https://doi.org/10.1111/j.1096-3642.1921.tb03288.x
- Subclass Opisthobranchia in British Columbia.https://ibis.geog.ubc.ca/biodiversity/efauna/documents/BCOpisthobranchsSandraMillen.pdf. Accessed on: 2021-2-06.
- A new species of Janolus Bergh, 1884 from the Northeastern Pacific.Spixiana39(1):15‑21.
- Phylum Ctenophora: list of all valid species names. http://faculty.washington.edu/cemills/Ctenolist.html. Accessed on: 2021-2-04.
- Large-scale impacts of sea star wasting disease (SSWD) on intertidal sea stars and implications for recovery.PLOS One13(3):e0192870. https://doi.org/10.1371/journal.pone.0192870
- Weather and climate. In:Compendium of forest hydrology and geomorphology in British Columbia. Land Management Handbook.Ministry of Forests and Range,Victoria.
- Mammals of British Columbia (2014). https://ibis.geog.ubc.ca/biodiversity/efauna/documents/MammalsofBCChecklist.pdf. Accessed on: 2021-2-13.
- The North and South American ascidians.Bulletin of the American Museum of Natural History84:1‑476.
- A second list of Bryozoa (Polyzoa) from the Vancouver Island Region.Contributions to Canadian Biology and Fisheries3:47‑131. https://doi.org/10.1139/f26-003
- A preliminary list of Bryozoa (Polyzoa) from the Vancouver Island Region.Contributions to Canadian Biology and Fisheries1:143‑201. https://doi.org/10.1139/f22-010
- Bryozoa of the Pacific coast of America. Part 1, Cheilostomata – Anasca.Allan Hancock Pacific Expedition14(1):1‑269.
- Bryozoa of the Pacific coast of America. Part 2, Cheilostomata – Ascophora.Allan Hancock Pacific Expedition14(2):271‑611.
- Bryozoa of the Pacific coast of America. Part 3, Cyclostomata, Ctenostomata, Entoprocta, and addenda.Allan Hancock Pacific Expedition14(3):613‑841.
- New species of Lissodendoryx Topsent, 1892 (Demospongiae, Poecilosclerida Coelosphaeridae) and Myxilla Schmidt, 1862 (Demospongiae, Poecilosclerida Myxillidae) from the Northeast Pacific.Zootaxa4700(1). https://doi.org/10.15468/dmunb4
- Food web complexity and species diversity.American Naturalist100(910):65‑75. https://doi.org/10.1086/282400
- Marine fishes of British Columbia (2002). https://ibis.geog.ubc.ca/biodiversity/efauna/documents/MarineFishofBC.pdf. Accessed on: 2021-2-13.
- Biodiversity citizen science: Outcomes for the participating citizens.People and Nature3(2):294‑311. https://doi.org/10.1002/pan3.10193
- Schizoporella unicornis — an alien bryozoan introduced into the Strait of Georgia.Journal of the Fisheries Research Board of Canada27:1847‑1853. https://doi.org/10.1139/f70-201
- QGIS Geographic Information System.Open Source Geospatial Foundation Project
- Delayed effects of capture-induced barotrauma on physical condition and behavioral competency of recompressed yelloweye rockfish, Sebastes ruberrimus.Fisheries Research186:258‑268. https://doi.org/10.1016/j.fishres.2016.09.004
- R: A language and environment for statistical computing.R Foundation for Statistical Computing,Vienna, Austria.
- Four new species of Hexactinellida (Porifera) and a name replacement from the NE Pacific.Zootaxa4466(1):124‑151. https://doi.org/10.11646/zootaxa.4466.1.11
- Why timing matters in a coastal sea: trends, variability and tipping points in the Strait of Georgia, Canada.Journal of Marine Systems131:36‑53. https://doi.org/10.1016/j.jmarsys.2013.11.003
- A new critical estimate of named species-level diversity of the recent Mollusca*.American Malacological Bulletin32(2). https://doi.org/10.4003/006.032.0204
- Natural history collection. http://search-collections.royalbcmuseum.bc.ca/. Accessed on: 2021-2-13.
- Invertebrate zoology: A functional evolutionary approach.7th ed.Thomson-Brooks/Cole,Belmont,1008pp.
- First occurrence of the non-native bryozoan Schizoporella japonica Ortmann (1890) in Western Europe.Zootaxa3780(3):481‑502. https://doi.org/10.11646/zootaxa.3780.3.3
- The genera Urticina and Cribrinopsis (Anthozoa: Actiniaria) from the north‐western Pacific.Journal of Natural History40:359‑393. https://doi.org/10.1080/00222930600703532
- Redescription of some species of Scionella Moore, 1903, with a review of the genus and comments on some species of Pista Malmgren, 1866 (Polychaeta: Terebellidae).Ophelia Supplemental5:239‑247.
- Evidence for a trophic cascade on rocky reefs following sea star mass mortality in British Columbia.PeerJ4:4:e1980. https://doi.org/10.7717/peerj.1980
- Climate variability and change of Mediterranean‐type climates.Journal of Climate32(10):2887‑2915. https://doi.org/10.1175/JCLI-D-18-0472.1
- Sea anemones of British Columbia. https://ibis.geog.ubc.ca/biodiversity/efauna/documents/SeaAnemonesofBC2010.pdf. Accessed on: 2021-2-04.
- Global diversity of Ascidiacea.PLOS One6(6):e20657. https://doi.org/10.1371/journal.pone.0020657
- Distribution of the Tanaidacea: synopsis of the known data and suggestions on possible distribution patterns.In:Crustacean biogeography.Routledge,Milton Park,316pp. https://doi.org/10.1201/9781315140674-8
- Galiano Island BC Canada Marine Zoology 1893–2021. Biodiversity Data Journal. Occurrence dataset. URL: https://doi.org/10.15468/gv9cy5
- Phylogenetic restriction of plant invasion in drought‐stressed environments: Implications for insect‐pollinated plant communities in water‐limited ecosystems.Ecology and Evolution11(15):10042‑10053. https://doi.org/10.1002/ece3.7776
- State of the Salish Sea.Salish Sea Institute, Western Washington University. https://doi.org/10.25710/vfhb-3a69
- Marine Ecoregions of the World: A bioregionalization of coastal and shelf areas.BioScience57(7):573‑583. https://doi.org/10.1641/b570707
- Nemertean taxonomy—Implementing changes in the higher ranks, dismissing Anopla and Enopla.Zoologica Scripta48(1):118‑119. https://doi.org/10.1111/zsc.12317
- Annelid phylogeny and the status of Sipuncula and Echiura.BMC Evolutionary Biology7(57):1‑11. https://doi.org/10.1186/1471-2148-7-1
- Phoronis ovalis (Phoronida, Lophophorata) in the White Sea: the first discovery of phoronids in the Arctic Basin.Doklady Biological Sciences374:523‑525.
- New data on distribution, morphology and taxonomy of phoronid larvae (Lophophorata: Phoronida).Invertebrate Zoology6(1):47‑64. https://doi.org/10.15298/invertzool.06.1.05
- Global change and local solutions: Tapping the unrealized potential of citizen science for biodiversity research.Biological Conservation181:236‑244. https://doi.org/10.1016/j.biocon.2014.10.021
- Brachiopod populations: distribution in fjords of British Columbia (Canada) and tolerance of low.Marine Ecology-Progress Series47:117‑128. https://doi.org/10.3354/meps047117
- Global diversity of sponges (Porifera).PLOS One7(4):e35105. https://doi.org/10.1371/journal.pone.0035105
- The marine zooplankton of British Columbia.Museum and Art Notes4(4):159‑165.
- Annotated checklist of the living sharks, batoids and chimaeras (Chondrichthyes) of the world, with a focus on biogeographical diversity.Journal of Fish Biology88(3):837‑1037. https://doi.org/10.1111/jfb.12874
- Darwin Core: An evolving community-developed biodiversity data standard.PLOS One7(1). https://doi.org/10.1371/journal.pone.0029715
- Distribution and abundance of marine mammals in the coastal waters of British Columbia, Canada.Journal of Cetacean Research and Management9:15‑28.
- World Register of Marine Species. https://www.marinespecies.org. Accessed on: 2021-1-12.
- Pacific coast pelagic invertebrates: a guide to the common gelatinous animals.2nd ed.Monterey Bay Aquarium,Monterey.
- Ascidians as models for studying invasion success.Marine Biology162(12):2449‑2470. https://doi.org/10.1007/s00227-015-2734-5
Supplementary materials
Formatted checklist summarising marine animal diversity reported for Galiano Island, British Columbia, Canada (1859–2021).
Curated taxonomic summary and catalogue of occurrence records documenting Porifera reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurence records documenting Cnidaria reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurrence records documenting Ctenophora reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurrence records documenting Nemertea reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurrence records documenting Platyhelminthes reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurrence data documenting Chaetognatha reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurrence records documenting Mollusca reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated summary and catalogue of occurrence records documenting Annelida reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated summary and catalogue of occurrence records documenting Sipuncula reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurrence records documenting Crustacea reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurrence records documenting Entoprocta reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurrence records documenting Brachiopoda reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurrence records documenting Bryozoa reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurrence records documenting Phoronida reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurrence records documenting Echinodermata reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurrence records documenting Tunicata reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurrence records documenting Actinopterygii reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurrence records documenting Chondrichthyes reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.
Curated taxonomic summary and catalogue of occurrence records documenting Mammalia reported for Galiano Island, British Columbia, Canada, in Darwin Core Standard format.